Portaalsite voor de échte zeeaquariaan
Zoeken op de site
Sponsors







Partners








Mysterieuze sluipmoordenaar onder de flowerkoralen (2008)
With speciall thanks to Eric Borneman for this articlle and pictures.
http://reefkeeping.com/issues/2008-01/eb/index.php
http://reefkeeping.com/issues/2008-01/eb/index.php
Vertaling: Ivan Baeten
Inleiding
Vanaf de late jaren tachtig en een eind in de jaren negentig behoorde het flowerkoraal (Catalaphyllia jardinei Wells 1971) tot de makkelijk houdbare koralen. Deze mooie dieren hebben grote groene fluorescerende poliepen met lange tentakels wat hen onderscheidt van de meeste andere koralen. De fluorescentie van Catalaphyllia onder actinisch of blauw licht is adembenemend. Helder groene en witte fijne strepen sieren de mondschijf, afgewisseld met contrasterende blauwe, oranje of purperen tentakeluiteinden. Het hoeft dan ook niet te verbazen dat dit begerenswaardige koralen zijn.
Catalaphyllia zijn normaal gewillige eters. De nematocysten in hun tentakels vertonen dezelfde plakkerige eigenschap zoals deze terug te vinden is bij anemonen. Hun weefsel is bijna volledig intrekbaar maar blijft ’s nachts meestal gedeeltelijk open staan. De mate waarin zij kunnen expanderen gekoppeld aan hun sterk netelende cellen maakt naburige koralen vatbaar voor serieuze beschadigingen. Hun eigen weefsel in volle expansie kan ook makkelijk beschadigd raken. Daarom is de nodige voorzichtigheid aan de orde wanneer een openstaande poliep uit het water gehaald dient te worden.
Als familielid van Euphyliidae is het skelet van Catalaphyllia bijna identiek aan de flabello-meandroid
(Benaming voor de skeletvorm waarbij ‘valleien’ gevormd worden zonder gemeenschappelijke wanden, red.) Euphyllia-soort. Zoals Trachyphyllia hecht Catalaphyllia zich vaak tijdelijk aan substraat als ze jong zijn. Eens volwassen maken ze zich meestal los van het substraat. Wanneer ze vast blijven zitten aan het rifsubstraat groeien ze uit tot grotere kolonies met een meandroid-vorm (skeletvorm waarbij ‘valleien’ gevormd worden met gemeenschappelijke wanden, red.). In het geval dat ze als losstaande koralen in zacht substraat leven, leidt dit tot een beperktere groei van de kolonies.
Figuur 1.
Deze koralen scheiden zich occasioneel af van hun skelet en dit bij slechte waterkwaliteit of andere vormen van stress (Figuur 1). Dit is geen normale vorm van reproductie maar blijkt een vluchtreactie te zijn die steevast leidt tot de dood van de kolonie. De losgekomen poliepkolonie kan in goed kwalitatief water en met de nodige zorg nog enige tijd blijven leven maar vormt hierbij geen skelet meer.

Figuur 2.

Figuur 3.
Figuur 3.
Aseksuele voortplanting is ongewoon. Kleine knopen, vergelijkbaar met deze van Euphyllia, ontstaan soms aan de rand van het skelet en kunnen wanneer ze loskomen van de moederkolonie nieuwe dieren vormen. Ouderkolonies kunnen in fragmenten van 2 centimeter worden gezaagd met een in zeewater gedompelde tegelzaag zonder veel risico op overlijden. Mijn koraal is gestart als een stek van 2 cm (Figuur 2). Het poliepweefsel is diep verankerd in het skelet. Het gebruik van hamer en beitel, tang, Dremel-gereedschap of andere groffe snijmethodes is normaliter zeer vernietigend voor het weefsel en resulteert in een lage overlevingskans voor de fragmenten.
Seksuele voortplanting is zeldzaam en haast niet gekend, zelfs niet in het wild. Kolonies zijn ofwel broedkolonies of hermafrodiete kuitschieters die hun zaad- en eicellen in pakketjes vrijlaten. Beide voortplantingsmethodes werden waargenomen in aquaria (Figuur 3) maar het is niet geweten waarom, waar en onder welke omstandigheden een bepaalde methode wordt gebruikt.
Catalaphyllia zijn terug te vinden in de Stille Zuidzee van Noord-Australië tot de Filippijnen en Indonesië, Zuidoost-Azië en ten zuiden van Japan. Je treft ze ook aan in het westelijk deel van de Indische Oceaan bij de Afrikaanse Oostkust. De soort komt algemeen tot overvloedig voor in afgeschermde zones met troebel water of lagunes met zachte of modderige bodems en tussen riffen waar ook zeegrassen te vinden zijn. Ze zijn niet algemeen verspreid maar komen geconcentreerd voor op bepaalde plekken waar ze dan ook veelvuldig aanwezig kunnen zijn.
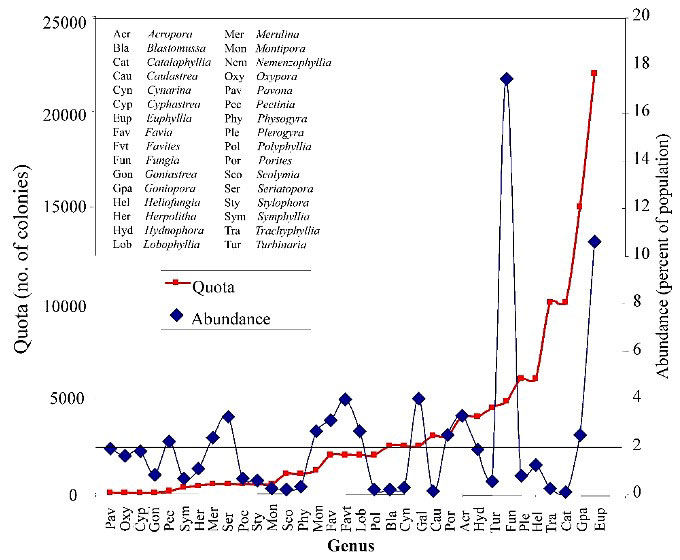
Bijna alle Catalaphyllia in de aquariumhandel zijn afkomstig van Indonesië. Een klein deel komt van enkele landen uit Zuidoost-Azië waar het verzamelen minder gereglementeerd is. Australië is recent ook gestart met de export. Indonesische oogstquota bedragen tienduizenden per jaar. De laatste jaren is dit getal gedaald waarbij het jaar per jaar varieert. In 2001 werd gestart met het bestuderen van de impact van de aquariumhandel op bepaalde soorten naar aanleiding van een EU-handelsstop.
Afgaand op de vindplaatsen is de reputatie van eenvoudig te houden koraal makkelijk te verklaren. Ze leven op dieptes van meer dan 40 meter, op ondiepe plaatsen met minder dan 1 meter waterstand, in troebel en helder water, vastgehecht aan rifsubstraat of losstaand in zachte bodem. Eind jaren negentig nochtans verschenen er dieren in de handel die het slachtoffer werden van een ziekte waarvan de oorzaak niet gekend was en waarvoor geen behandeling bestond. Na een tijd vertoonden bijna alle exemplaren tekenen van deze dodelijke aandoening wat resulteert in een opgezwollen mondschijf met ingetrokken tentakels aan de rand (Figuur 4). In vele gevallen is er een witte ondoorschijnende slijmerig web aanwezig op de mondschijf (Figuur 5). Reacties bij aanwezigheid van voedsel nemen af en de ‘plakkerigheid’ van de tentakels is verdwenen waardoor zelfs het grijpen van onbeweeglijke voedseldeeltjes niet meer mogelijk is. Het koraalweefsel krimpt uiteindelijk in en de kolonie sterft ondanks alle mogelijke experimenten welke ondertussen zijn uitgeprobeerd (Figuren 6.1 en 6.2). Meerdere auteurs binnen de aquariumhobby hebben over dit verschijnsel geschreven en theorieën voorgesteld die een verklaring zouden geven waarom deze dieren momenteel niet overleven. Op internetfora en in tijdschriften werd reeds stevig gespeculeerd waarom de koralen niet langer gedijen in aquariums. Geen van deze speculaties heeft echter geleid tot een beter begrip van het fenomeen. Omdat deze kwaal alle tekenen vertoont van een ziekte zal ik ernaar refereren als het flowerkoraalsyndroom (FKS) en dit totdat een geschiktere benaming kan gegeven worden op basis van de ziekteverschijnselen. Ik heb nooit gehoord dat C. jardinei FKS heeft opgelopen zonder in direct of indirect contact te hebben gestaan met een ander exemplaar met FKS. Ondanks dat deze dieren kunnen sterven wegens verschillende oorzaken, C. jardinei is een sterke soort in gevangenschap waar geen FKS aanwezig is. Jaren geleden startte ik een project om de redenen te achterhalen omtrent de hoge sterftecijfers in de aquariumhandel en dit gekoppeld aan veldwerk en andere rapporteringmethodes.

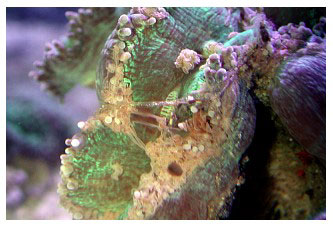
Figuur 4. Figuur 5


Figuur 6.1 Figuur 6.2
Figuur 4. Figuur 5
Figuur 6.1 Figuur 6.2
Materiaal en methodes.
Locaties
Het onderzoek werd uitgevoerd op 11 verzamelsites doorheen de Spermonde-archipel waarvan vermeld werd dat C. jardinei er voorkwam en waar tevens de meeste Catalaphyllia werden geoogst. Verschillende methodes waaronder snorkelen en duiken werden gebruikt om de populaties te beoordelen. Gesprekken werden gevoerd met de mensen die de koralen verzamelden, exporteurs en leden van de handelsgroep AKKII. Verzamelde koralen werden geteld, onderzocht en gemeten in zowel plaatselijke vangstations als verschillende grote exportbedrijven waarbij elk exemplaar met FKS werd gedocumenteerd.
Rapportering
Contacten werden gelegd met duikers, wetenschappers, verzamelaars, aquaristen en handelaars in Vietnam, India, Fiji, Australië en de Filippijnen. Rapporten werden tevens geschreven door aquaristen in Europa via internetfora en tijdens meerdere van mijn reizen doorheen verschillende Europese landen. Het initiële doel van de interviews en rapporten was om te bepalen of en in welke mate Catalaphyllia geïmpacteerd was door FKS in het wild en in aquaria. Ik wilde ook achterhalen in welke graad deze koralen met succes werden gehouden in aquaria en dat met betrekking tot die kolonies die tekenen van FKS vertoonden.
Staalnames
Catalaphyllia-exemplaren gebruikt voor het project kwamen vanuit verschillende hoeken, zowel gezonde als zieke dieren. Ik kreeg onder andere koralen met FKS van aquaristen. In totaal werden 79 exemplaren verzameld waarvan 70 met FKS of een andere aandoening. Zeven dieren leken gezond en vertoonden geen FKS en twee FKS-vrije exemplaren kwamen uit een aquarium waar ze ondertussen vijf jaar in hadden verbleven. 75 FKS getroffen kolonies werden gebruikt voor histologische analyse (weefselleer, red.) terwijl twee fragmenten van een gezonde kolonie als controle dienst deden. Ik slaagde er niet in om andere gezonde kolonies te krijgen als referentie (zie Discussie).
Ziektebeeld
Vijf exemplaren van C. jardinei met de eerste tekenen van FKS werden in een quarantainebak geplaatst en meerdere weken geobserveerd. Wijzigingen in de progressie van de ziekte werden genoteerd. Alvorens complete weefselontbinding optrad, werden de dieren gefixeerd zoals verder beschreven.
Experiment in functie van besmetting
Vijf exemplaren zonder FKS-aandoeningen werden in de buurt geplaatst van een ziek dier om na te gaan of de koralen FKS konden krijgen zonder direct contact. Deze gezonde dieren werden besteld bij een Indonesische verzamelaar (twee afkomstig van Jakarta, twee van Sulawesi en één van Sumatra). Ze kwamen niet in contact met andere Catalaphyllia of aquariumwater en verbleven enkel in het water van de transportzak tijdens de overbrenging. In een tweede proefopstelling werd een ziek exemplaar ondergebracht in een multi-aquariumsysteem waarvan het water gedeeld werd met een gezond dier dat zich in een ander aquarium bevond. Andere koraalsoorten die in deze aquaria stonden werden eveneens geobserveerd op tekenen van stress of ziekte veroorzaakt door de FKS-koralen.
Voorbereiding
Elk levend dier werd bij aankomst gefotografeerd, in een aquarium geplaatst en opnieuw gefotografeerd na acclimatisatie en weefselexpansie. Wanneer een zeer ziek koraal werd aangetroffen dan werden foto’s genomen gevolgd door fixatie voor histologiedoeleinden. Omdat de poliepen zich volledig kunnen terugtrekken in hun skelet was het noodzakelijk om de dieren voldoende tijd te geven in het aquarium om zich te openen zodat hun toestand kon beoordeeld worden. Wanneer het ziektebeeld overeenstemde met FKS werd het koraal verwijderd uit het aquarium en gefixeerd. Het aquariumwater werd gesteriliseerd door toevoeging van bleekwater waarna het mengsel werd afgevoerd naar de waterzuiveringsinstallatie. Dit werd niet toegepast in het besmettingsexperiment waar alle water continu doorheen het hele systeem werd gepompt. Sommige exemplaren werden gefixeerd voor transport nadat de afzender ze had gefotografeerd en FKS had vastgesteld. Fixatie gebeurde aan de hand van vooraf beschreven protocollen. Afhankelijk van het fixeermiddel (alcohol of formol) werden de koralen opgeslagen of nabehandeld met zinkformaline. Levende koralen werden twee tot drie dagen gefixeerd afhankelijk van hun grootte en hoeveelheid weefsel en dit in zinkformaline. Hierna werden ze ontkalkt totdat alle kalkskelet was verwijderd. Nota’s werden genomen wanneer abnormaal organisch materiaal of organismen overbleven na ontkalking zoals Lithophaga sp., borstelwormen, sponzen, algen en schimmelfilamenten. Fracties van de ontkalkingsvloeistof werden microscopisch onderzicht op micro-organismen die overeenkomen met FKS-koralen. Ontklakte weefsels werden gespoeld in gedemineraliseerd water en vervolgens in 70 % ethanol gestoken voor opslag en transport. De koralen werden geregistreerd en samen met de benodigde formulieren opgestuurd naar het International Registry of Coral Pathology (IRCP) voor histologisch onderzoek. Totaalfoto’s van de weefsels werden genomen waarna de koralen in verschillende richtingen werden versneden tot plakjes van 6 µm. De plakjes werden op glasplaatjes gelegd en gekleurd voor weefselonderzoek. De plakjes werden onderzocht onder drie verschillende types van microscopen om een algemeen beeld te kunnen vormen van het weefsel en om mogelijke etiologische reagens aan te treffen (etiologie is dat onderdeel van de pathologie dat zich bezighoudt met de oorzaken van ziekten, red.).
Resultaten
Locaties
In alle geografisch onderzochte gebieden kwam Catalaphyllia jardinei zelden voor, namelijk tussen 0.03 en 0.05 exemplaren per vierkante meter. Ze kwamen voor in vastgehechte en losstaande vorm in oppervlakkig en diep water, in verslibte gebieden en op zachte bodems. Tijdens ons verblijf bij de Indonesische verzamelaars van de grote verzamelgebieden van Makassar, Sulawesi, werd ons verzekerd dat Catalaphyllia algemeen voorkwam en dat ze geoogst werden op meerdere plaatsen. Echter, de koralen waren zeldzaam op alle sites welke we bezochten en de aantallen opgegeven door de verzamelaars overstegen steeds in grote mate de hoeveelheden die we effectief aantroffen. We vonden kolonies op meerdere verschillende locaties en net zoals Euphyllia in sterk uiteenlopende habitats. Op grotere dieptes (> 40 m) waar de meest kleurrijke rode en groene Trachyphyllia werden verzameld, werden ook de meeste Catalaphyllia geoogst. Dit is een zeer lichtschuwe omgeving met een zand- en slibbodem zonder hard substraat. Het is dun bevolkt met losstaande koralen, macro-algen en cyanobacteriën. Hier zagen we Catalaphyllia als kleien losstaande kolonies waarvan de grootte en vorm overeenstemt met deze die je aantreft in de aquariumwinkels. Alle exemplaren afkomstig uit dit gebied hadden purperen tentakeluiteinden. Geen enkel exemplaar van elk ander gebied dat we bezochten vertoonde deze begerenswaardige kleur. Veron (2000) schrijft echter dat de aanwezigheid van gekleurde tentakeluiteinden een veel voorkomend kleurpatroon is. Tegengesteld waren de weinige kolonies die we vonden op geïsoleerde en ondiepe riffen (‘patch reefs’). Daar groeiden de koralen op een hard substraat en waren ze veel groter met een uitgesproken flabello-meandroid vorm. De mondschijf was groen en wit gestreept en de tentakels vertoonden geen gekleurde punten. Deze kolonies worden verwijderd met hamer en beitel of met een koevoet waardoor de spierwitte skeletbasis beschadigd wordt. Dieren van andere sites hebben een verkleurde en conisch gevormde skeletbasis die typisch is voor losstaande kolonies. Een ander verzamelgebied was een ondiepe zone vlakbij de kust waar saai gekleurde Trachyphyllia werden geoogst. Hiervan werd beweerd dat het een uitstekende locatie voor het verzamelen van Catalaphyllia was. De site bleek echter verstoken van kolonies. Verklaringen hiervoor zouden kunnen zijn dat de dieren ooit sporadisch zijn voorgekomen, een lage natuurlijke bevolkingsgraad of leeg geoogst. Vergelijkbaar met de diepwaterhabitat, leefden de kolonies hier als losstaande exemplaren en nooit vastgehecht. Ze waren bijna zo goed als begraven in fijn slib en klein in hun soort vanwege het substraat waar ze in zaten. Vanwege dit slib had hun skelet een smerig bruine kleur. Het weefsel was vaalbruin met groenbruine mondschijven en tentakels.
Er werd ook een gebied beschreven met zeegrasbegroeiing waar Catalaphyllia veelvuldig zou voorkomen maar gelijkaardige gebieden die wij bezochten, bleken raar of zelden exemplaren te huisvesten. Sites met zachte bodem waar de meeste kolonies werden aangetroffen, leefde de soort losstaand met een dichtheid van 0.05 kolonies per vierkante meter. Het geïsoleerde en ondiepe rif bevatte voornamelijk grote aangehechte kolonies met meandroid-groei waarvan de exemplaren te groot waren voor de handel. Wetende dat landen nog steeds Catalaphyllia waarnemen in regio’s waar geen dieren worden gevangen voor de handel terwijl ze niet langer te vinden zijn in Indonesische vanggebieden, lijkt overbevanging een logische verklaring voor de lage aanwezigheid in de gebieden waarvan men beweert dat Catalaphyllia veelvuldig aanwezig zouden moeten zijn.
Figuur 7.
Figuur 7.
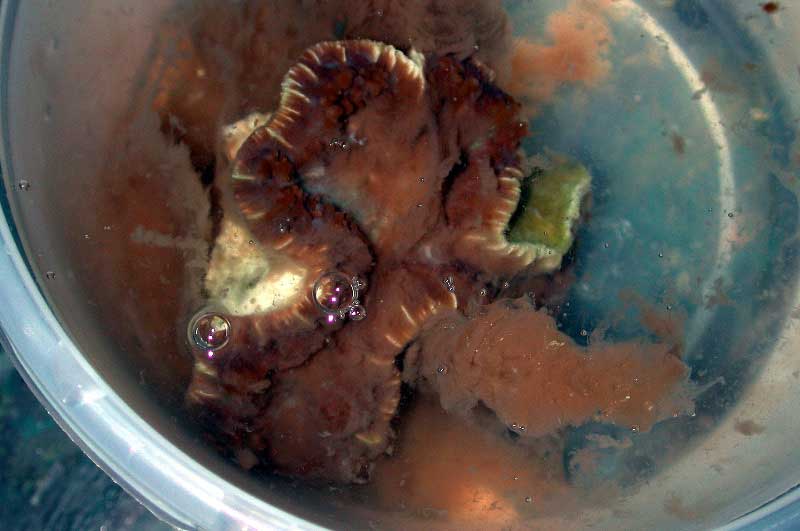
In geen enkele situatie troffen we FKS-dieren aan in het wild. Ditzelfde geldt voor de dieren die onderzocht werden in de vangstations, de plaats waar ze bewaard worden alvorens te verhuizen naar de exportfirma’s (Figuur 7). Meerdere grote exporteurs hielden C. jardinei in grote betonnen vijvers waarin honderden exemplaren staan. Door de weinige ruimte was er veel contact tussen de verschillende dieren. Het beschikbare water per exemplaar was laag en waterwissels gebeurde wekelijks met door tankwagens aangevoerd zeewater. Bij één exporteur werd één ziek exemplaar aangetroffen (Figuur 8).


Figuur 8. Figuur 9.
Figuur 8. Figuur 9.
Rapportering
Via interviews met mensen uit landen waar Catalaphyllia wordt gevonden en gehouden in aquaria bleek dat niemand ooit een FKS-exemplaar had gezien in het wild of in aquaria. Dit was opvallend voor Australië waar de koralen algemeen voorkomen, het verzamelen wordt gecontroleerd en er veel aquaristen zijn. Even bijzonder is dat Australië de import vanuit andere regio’s verbiedt. In Europa werden wel gevallen van FKS gerapporteerd maar de cijfers lagen lager dan die van de US. Europa ontvangt hun koralen van dezelfde gebieden als de US en Indonesië is de voornaamste exporteur van Catalaphyllia voor beide. De grootste hoeveelheid gaat naar de US.
Staalnames
Ondanks een zorgvuldig beschreven protocol om de dieren te verzamelen, behandelen en transporteren, ontving ik vele slechte zendingen. In sommige gevallen werd het protocol onvoldoende gevolgd, lekten transportzakken en werden fixeermiddelen niet correct toegepast. Nabehandeling werd uitgevoerd om de fixatie te bevorderen voor het histologisch onderzoek in zo’n gevallen. In meerdere zendingen stierven de dieren tijdens transport; deze werden uit de studie gehaald (Figuur 9). Andere vormen van staalnames zoals uitstrijkjes en foto’s en documentatie werden niet geleverd. De exemplaren die ik verzamelde werden met de nodige voorzichtigheid behandeld en verzorgd alvorens ze werden gefixeerd met zinkformol (werd tevens gebruikt voor postfixatie). Het gebruik van verschillende andere fixeermiddelen in bepaalde zendingen werd vastgesteld.
Experiment rond besmettingsgevaar
Experiment 1
Vijf koralen die individueel werden gevangen en verpakt, werden opgestuurd naar een koraalkwekerij in Houston. Ze leken allemaal gezond bij aankomst. Drie stuks werden in mekaars buurt gezet zonder mekaar te raken en dit in een systeem met meerdere koraalsoorten en met elkaar verbonden aquaria. Dit systeem had nog nooit FKS-koralen gehuisvest. Eén exemplaar (Jakarta 1) werd in een apart en kleiner aquarium geplaatst waarin zich andere koralen bevonden naast vissen en ongewervelden maar waar nooit een C. jardinei had in gezeten. De laatste (Sulawesi 2) werd in een iets groter maar gelijkaardig aquarium met een mix aan dieren (Figuur 10). Geen enkel koraal in de verschillende aquaria vertoonde enig teken van ziekte, leken allemaal gezond en groeiden. De waterkwaliteit in alle aquaria was excellent en vergelijkbaar met dit van oceaanwater. Licht varieerde van 800 tot 60 PAR (indicatie voor de hoeveelheid fotosynthetische straling) wat ruwweg overkomt met de natuurlijke condities in het wild.
Figuur 10.
&
Figuur 10.
&
Schijfanemonen van het Actinodiscus type
Systematiek:
Orde: Corallimorpharia
Familie: Discosomatidae
Genus: Discosoma (Actinodiscus)
Amplexidiscus
Rhodactis
Metarhodactis

Foto Hans Peter, Discosoma inchoata.
Algemeen:
Actinodiscus of Discosoma schijfanemonen worden al zolang gehouden als er zeewateraquaria bestaan. We noemen ze schijfanemonen doch eigenlijk zijn het zachte koralen. Het zijn waarschijnlijk de gemakkelijkst te vinden zachte koralen, alhoewel een mooi assortiment samenstellen niet zo gemakkelijk blijkt te zijn.
Purperen en rode soorten zijn overal verkrijgbaar, andere kleuren minder.
Ze hebben geen exoskelet maar groeien op rotsen en stenen.
De naam is afgeleid van hun verschijningsvorm, namelijk een korte steel met een schijf bovenop. Deze schijf kan volledig rond zijn of gerafeld. Gewoonlijk is de korte steel niet te zien.
Schijfanemonen kunnen enkel- of veelkleurig zijn, glad of ruw, met knobbels en strepen die vanuit het midden naar de rand toelopen.
Schijfanemonen zijn over het algemeen “harde” aquariumdieren, wat niet wegneemt dat ze “mishandeling” voor lief nemen.
Wat men bedoeld is dat schijfanemonen kunnen leven in water van verschillende samenstelling, wat ze niet verdragen is plotse veranderingen in deze samenstelling.

Foto Hans Peter, Discosoma inchoata "Tonga Blue"
Plotse veranderingen in de waterhuishouding is slecht voor ALLE dieren en lagere dieren in een aquarium. Het is een primaire oorzaak voor stress, en stress veroorzaakt ziekte en aanvallen van parasieten door verzwakte weerstand.
Veranderingen die we zeker moeten vermijden zijn plotse temperatuurwijzigingen, abrupte schommelingen in pH of zoutgehalte.
Schijfanemonen komen niet alleen in verschillende groeiwijzen voor doch ook in verschillende kleuren. Dit heeft te maken met de omgeving waar ze voorkomen en welk soort en intensiteit van het licht ze daar ontvingen.
Dit bepaalt hun pigmentering en welke kleur zal domineren.
Sommige soorten zijn egaal gekleurd, andere hebben verschillende kleuren.

Foto Hans Peter, Rhodactis sp.
Het moet gezegd; vele schijfanemoonsoorten zijn niet bij naam beschreven en de meeste soorten worden als”Actinodiscus sp.” Voorgesteld.
Het is waarschijnlijk de groep dieren die het minst door wetenschappers is onderzocht. Misschien weten sommige aquarianen er meer van dan de wetenschap!
Bovenop de verschillen in vorm en kleur komen nog eens de verschillen in textuur van de poliepen, ze kunnen zeer glad tot knobbelvormig zijn soms met kleine tentakeltjes. De variatie is oneindig. Deze grote variatie in vormen en kleuren maken deze zachte koralen zo interessant voor het zeeaquarium.
Schijfanemonen komen voor in alle oceanen waar riffen te vinden zijn. In de vrije natuur komen ze vooral voor in dichte clusters. In het aquarium kan men dit ook gemakkelijk bereiken (knopvorming).
Om dit te verwezenlijken moet de waterkwaliteit parameters bezitten die verder in dit artikel voorgesteld worden.
Het is praktisch onmogelijk om individuele specimen te vinden die niet tot een kolonie behoren.

Foto Hans Peter,
In het aquarium:
Het zijn waarschijnlijk de “hardste” zachte koralen die we kunnen vinden, al geloven sommige aquarianen dat niet altijd omdat ze in het verleden problemen hebben gehad met deze lagere dieren. Deze problemen zijn in hoofdzaak te wijten aan de waterkwaliteit of verkeerde belichting of stroming.
Niet alle actinodiscussoorten vallen in dezelfde behoefte categorie.
Aquarianen moeten experimenteren om te ondervinden wat de beste verlichting en de beste stroming is voor de soort die ze hebben verworven.
De meeste schijfanemonen doen het goed bij een medium tot sterke belichting. Ideaal echter zou zijn wanneer de reactie kan geobserveerd worden onder verschillende intensiteiten van verlichting. Hetzelfde voor de soort en richting van de stroming die ze ontvangen.
Sommige variëteiten houden van sterke stroming, andere meer gematigde stroming, er is geen allesomvattende aanbeveling voorhanden die voor alle soorten geldt.
Observeren hoe verschillende specimen reageren op wisselende condities kan men best bereiken door ze gedurende een à twee weken aan deze parameters bloot te stellen en dan met intervallen foto’s nemen om de evolutie te vergelijken.
Voorgestelde waterparameters:
Temperatuur: 25 tot 27 º C.
Zoutgehalte: S.G.: 1.022 – 1.024
pH : 8.1 – 8.3
Calcium: onbelangrijk, doch wanneer steenkoralen aanwezig +/- 450 ppm
Nitraten: minder dan 10 ppm
Fosfaat: minder dan 0.04 ppm
Silicaat: minder dan 0.5 ppm
Opgeloste zuurstof : 7 of hoger
Dagelijkse toediening van Jodium (aanwijzingen van fabrikant volgen)
Stroming: gemiddelde tot sterke stroming, NIET direct op de schijfanemonen richten.
Houdt borstelwormen uit het aquarium.
Heremietkreeften mogen niet over de schijfanemonen kruipen.
Er mogen geen algen op of tussen de individuele poliepen van de kolonie groeien.
Wanneer dit het geval is dan zijn er waarschijnlijk te hoge fosfaat- of silicaatwaarden.
J.R.
Het complexe koraal: immuniteit en energietransport
Koralen worden door velen nog steeds beschouwd als simpele organismen. De vrij eenvoudige lichaamsstructuur en lage rang op de evolutionaire ladder zijn hiervoor verantwoordelijk. Toch is dit beeld niet helemaal terecht. Zeer recente onderzoeken tonen aan dat koralen veel eigenschappen vertonen welke lijken op die van hoger ontwikkelde dieren. Op genetisch vlak zijn er veel overeenkomsten met gewervelde dieren zoals zoogdieren en zelfs mensen. Het is nu ook duidelijk geworden dat koralen een immuunsysteem hebben ontwikkeld; een interessant gegeven voor de aquariaan.
Stylophora pistillata
Wetenschappers beweren al jaren dat koralen niet onderschat moeten worden. Afgelopen jaar publiceerden wetenschappers van het Oceanografisch en Limnologisch Instituut te Eilat, Israël dat de steenkoraalsoort Stylophora pistillata in staat is vreemd van eigen weefsel te onderscheiden1.
 Dit onderzoek werd uitgevoerd als onderdeel van het CORALZOO project. Zowel de mogelijke groeivormen als het gedrag van dicht bij elkaar geplaatste kolonies werd bepaald. De experimenten werden verder in situ (in de natuurlijke omgeving) uitgevoerd. Zij plaatsten zeer jonge S. pistillata kolonies dicht naast elkaar op het rif; genetisch identieke fragmenten (afkomstig van dezelfde moederkolonie) en genetisch verschillende. De leeftijd van de kolonies varieerde tussen de 0 (net na metamorfose van de koraallarven naar primaire kolonies) en 4 maanden. Al na enkele dagen waren de eerste resultaten zichtbaar; deze omvatten fuseringen en afstotingen. De gefuseerde kolonies werden chimeren genoemd; dit zijn organismen met genetisch verschillende weefsels (naar het bekende Griekse mythologische wezen; een fusie tussen verschillende dieren).
Dit onderzoek werd uitgevoerd als onderdeel van het CORALZOO project. Zowel de mogelijke groeivormen als het gedrag van dicht bij elkaar geplaatste kolonies werd bepaald. De experimenten werden verder in situ (in de natuurlijke omgeving) uitgevoerd. Zij plaatsten zeer jonge S. pistillata kolonies dicht naast elkaar op het rif; genetisch identieke fragmenten (afkomstig van dezelfde moederkolonie) en genetisch verschillende. De leeftijd van de kolonies varieerde tussen de 0 (net na metamorfose van de koraallarven naar primaire kolonies) en 4 maanden. Al na enkele dagen waren de eerste resultaten zichtbaar; deze omvatten fuseringen en afstotingen. De gefuseerde kolonies werden chimeren genoemd; dit zijn organismen met genetisch verschillende weefsels (naar het bekende Griekse mythologische wezen; een fusie tussen verschillende dieren).
De chimeren bestonden uit 2, 3 of zelfs 6 verschillende kolonies (figuur 1). Chimeren bestaande uit 4 of meer genetisch verschillende kolonies werden multi-partner entities (meerdere-partners entiteit, of MPE) genoemd. Verder zag men 2 of 3 kolonies welke elkaar tegelijk afstootten. Na fusie was bijna niet meer te zien wat de oorspronkelijke kolonies waren, hoewel niet alle fusies permanent waren. De afstotingsverschijnselen waren divers; afwezigheid van weefselverbinding waarbij skeletten elkaar raakten, overgroei van één van de kolonies, bleking, necrose (afsterving) en verder dode kolonies. Aanvankelijk kwamen fusie- en afstotingsverschijnselen even vaak voor, maar naarmate de chimeren uit meer partners bestonden nam dit laatste af. Hoe groter de chimeer, hoe kleiner de kans dat een nieuwe kolonie werd afgestoten.
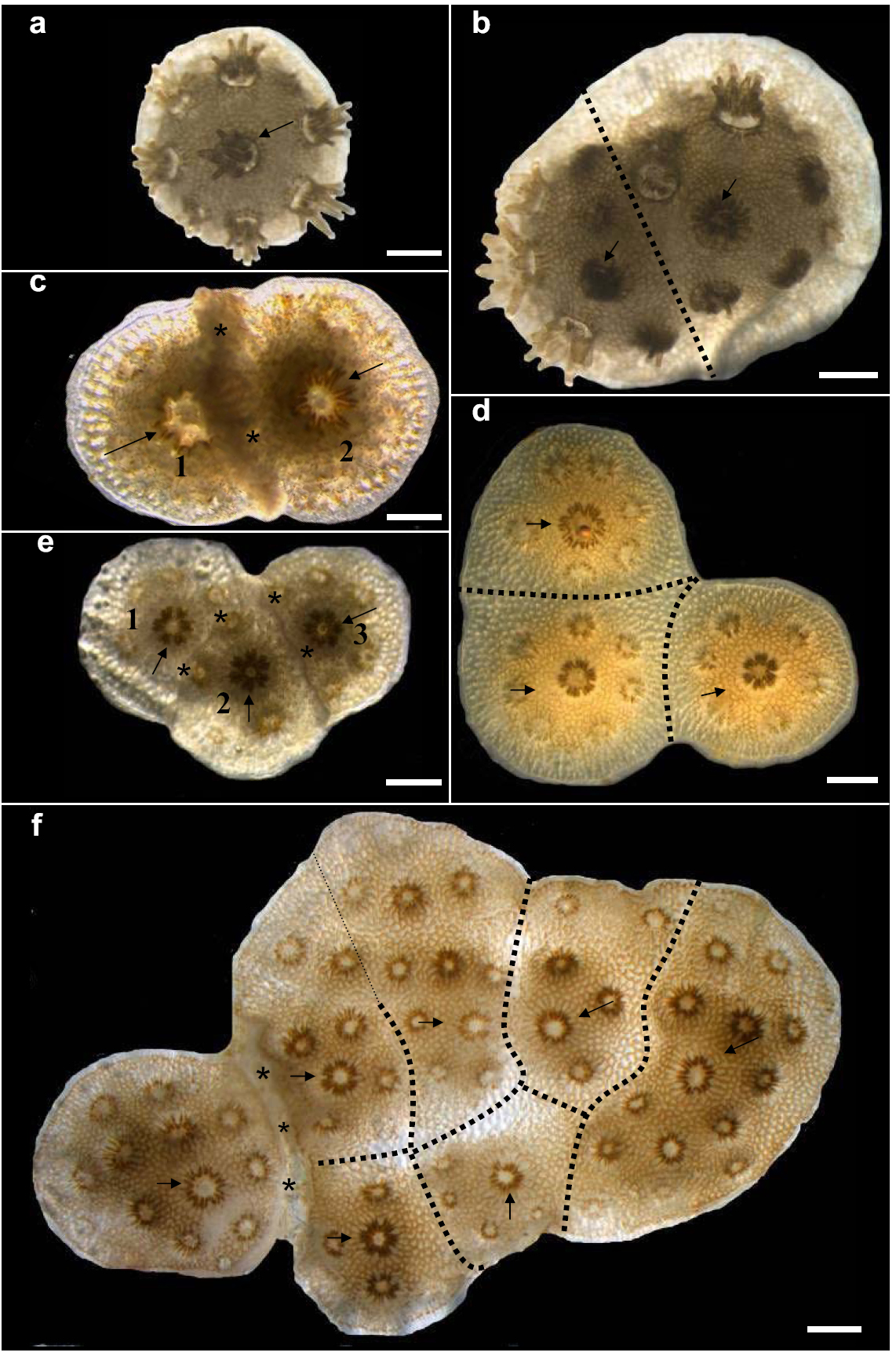 Figuur 1: Stylophora pistillata kolonies van 0 tot 4 maanden oud, op zeer korte afstand naast elkaar geplaatst, vormden chimeren of stootten elkaar af. a: enkel genotype, 2 maanden oud. b: bi-chimeer; 2 gefuseerde kolonies c: 2 genotypen welke elkaar afstootten, d:tri-chimeer, 1,5 maand oud, e: 3 elkaar afstotende genotypen, f: samengestelde kolonie van 7 genotypen; een zogenaamde multi-partner entity (meerdere-partners entity of MPE). Opvallend is dat slechts 1 genotype wordt afgestoten, geheel links. Asteriksen geven afstoting aan, stippellijnen geven grenzen tussen oorspronkelijke kolonies aan, pijlen geven de primaire poliepen (poliepen waaruit de kolonie is ontstaan, na metamorfose) aan. De witte schaalbalken rechts onderaan zijn 1 mm (Shaish et al, PloS ONE, 2008).
Figuur 1: Stylophora pistillata kolonies van 0 tot 4 maanden oud, op zeer korte afstand naast elkaar geplaatst, vormden chimeren of stootten elkaar af. a: enkel genotype, 2 maanden oud. b: bi-chimeer; 2 gefuseerde kolonies c: 2 genotypen welke elkaar afstootten, d:tri-chimeer, 1,5 maand oud, e: 3 elkaar afstotende genotypen, f: samengestelde kolonie van 7 genotypen; een zogenaamde multi-partner entity (meerdere-partners entity of MPE). Opvallend is dat slechts 1 genotype wordt afgestoten, geheel links. Asteriksen geven afstoting aan, stippellijnen geven grenzen tussen oorspronkelijke kolonies aan, pijlen geven de primaire poliepen (poliepen waaruit de kolonie is ontstaan, na metamorfose) aan. De witte schaalbalken rechts onderaan zijn 1 mm (Shaish et al, PloS ONE, 2008).
Deze resultaten zijn heel interessant, aangezien de wetenschappers eerdere onderzoeken naar immuniteit bij koralen hiermee bevestigden. Koralen hebben duidelijk een subtiele manier gevonden om vreemd van eigen weefsel te onderscheiden; een sterk kenmerk van een immuunsysteem. Verder kan worden geconcludeerd dat deze immuniteit is aangeboren, zoals bij veel dieren het geval is. Dit is zo omdat deze koralen nog niet eerder in contact waren gekomen met vreemd weefsel. In dit geval noemt men deze reacties op vreemd weefsel een allogene reactie; dit betekent dat het koraal reageert op genetisch verschillend weefsel van dezelfde soort. Het tegengestelde is isogeen; genetisch identiek, zoals een transplantatie van een orgaan tussen een één-eiige tweeling, en autogeen; zoals een transplantatie van het eigen weefsel (bijvoorbeeld huid) naar een andere locatie.
Zodra planula larven van deze soort zich op het rif vestigen, dan vindt in feite bovenstaand experiment plaats. De koralen reageren op elkaars weefsel, en dit zal zowel intra- als interspecifiek plaatsvinden (interacties tussen de eigen soort en andere soorten). Uiteindelijk zullen zich zowel solitaire kolonies als chimeren gaan vormen, hoewel het laatste een zeldzamer geval lijkt. Verder werd gevonden dat de gemiddelde grootte van de MPE- (4 of meer gefuseerde kolonies) leden kleiner was dan die van de solitaire kolonies (hoewel de MPE’s in totaal uiteraard wel groter waren). Dit is waarschijnlijk het resultaat van de interacties tussen de kolonies, zoals afstoting. Dit verbruikt veel energie, wat ten koste gaat van de groei. Het feit dat de MPE’s groter waren is een indicatie voor verhoogde overlevingskansen. De les die hieruit kan worden getrokken is dat hoewel een verbond tussen organismen voordelen heeft, er hiernaast ook nadelen aan kleven. Als er inderdaad kosten zijn zoals groei-inhibitie (remming) en zelfs sterfte, waarom vormen koralen dan toch deze MPE’s, of “superkolonies”? Overleven op het rif draait allemaal om het innemen van ruimte, en bij MPE’s werd gevonden dat deze sneller groeiden. Een snellere groei betekent het sneller innemen van een niche, een specifieke plaats, op het rif. Dit verhoogt vervolgens de overlevingskansen van deze kolonies, omdat zij binnen korte tijd een plek veroverd hebben waar zij licht kunnen opvangen en nutriënten kunnen opnemen.
 Deze resultaten laten zien dat koralen absoluut bijzondere dieren zijn. Regelmatig worden nieuwe manieren gevonden waarop koraalpoliepen samenwerken. Niet alleen kloneren (delen) veel soorten zich waarbij ze grote kolonies vormen, ze fuseren ook met genetisch verschillende individuen om hun overlevingskansen te vergroten. Dit verschijnsel, natuurlijk chimerisme, is ook waargenomen bij veel andere sessiele (vastzittende) mariene organismen. Voorbeelden hiervan zijn sponzen, zachte koralen, zakpijpen en zelfs bepaalde steenkoralen.
Deze resultaten laten zien dat koralen absoluut bijzondere dieren zijn. Regelmatig worden nieuwe manieren gevonden waarop koraalpoliepen samenwerken. Niet alleen kloneren (delen) veel soorten zich waarbij ze grote kolonies vormen, ze fuseren ook met genetisch verschillende individuen om hun overlevingskansen te vergroten. Dit verschijnsel, natuurlijk chimerisme, is ook waargenomen bij veel andere sessiele (vastzittende) mariene organismen. Voorbeelden hiervan zijn sponzen, zachte koralen, zakpijpen en zelfs bepaalde steenkoralen.
Lobophyllia corymbosa
Naast Stylophora pistillata, is ook bij andere koraalsoorten immuniteit ontdekt, waaronder bij de steenkoraalsoort Lobophyllia corymbosa2. Wetenschappers van de Universiteit van Tel Aviv en de nationale Universiteit van Ierland ontdekten recentelijk dat ook poliepen van L. corymbosa vreemd van eigen weefsel kunnen onderscheiden. Verder zagen zij ook dat deze poliepen nutriënten transporteerden naar verwonde exemplaren binnen de eigen kolonie.
De soort L. corymbosa lijkt wat op soorten van het genus Caulastrea; wanneer de kolonie volgroeid is, snoeren de poliepen hun weefsel volledig af waardoor deze solitair worden. De onderzoekers verwijderden 52 poliepen van 14 verschillende kolonies te Eilat, Israël. De poliepen werden of intact gelaten, of in de lengte vanaf boven doorgesneden. Vervolgens werden de poliepen, of helften daarvan, op verschillende manieren aan elkaar verbonden met koperdraad. Dit leverde 3 vormen van transplantatie op; autotransplantaties (twee poliephelften van de originele poliep), isotransplantaties (twee poliephelften van verschillende poliepen, maar van dezelfde kolonie, welke dus genetisch identiek zijn) en allotransplantaties (twee helften van genetisch verschillende poliepen).

De resultaten van de experimenten werden na 6 weken bekeken, waarbij verschillende aspecten werden gedocumenteerd; weefselfusie, skeletfusie, cytotoxische afstoot (productie van giftige stoffen), weefselovergroei en absentie van respons. Zij vonden dat geen van alle intacte poliepen een reactie vertoonde, zelfs niet wanneer twee intacte poliepen afkomstig van verschillende kolonies (dus genetisch verschillend) aan elkaar gebonden waren. Echter, de transplantaties van de poliephelften lieten iets heel anders zien.Stylophora pistillata
Wetenschappers beweren al jaren dat koralen niet onderschat moeten worden. Afgelopen jaar publiceerden wetenschappers van het Oceanografisch en Limnologisch Instituut te Eilat, Israël dat de steenkoraalsoort Stylophora pistillata in staat is vreemd van eigen weefsel te onderscheiden1.
Dit onderzoek werd uitgevoerd als onderdeel van het CORALZOO project. Zowel de mogelijke groeivormen als het gedrag van dicht bij elkaar geplaatste kolonies werd bepaald. De experimenten werden verder in situ (in de natuurlijke omgeving) uitgevoerd. Zij plaatsten zeer jonge S. pistillata kolonies dicht naast elkaar op het rif; genetisch identieke fragmenten (afkomstig van dezelfde moederkolonie) en genetisch verschillende. De leeftijd van de kolonies varieerde tussen de 0 (net na metamorfose van de koraallarven naar primaire kolonies) en 4 maanden. Al na enkele dagen waren de eerste resultaten zichtbaar; deze omvatten fuseringen en afstotingen. De gefuseerde kolonies werden chimeren genoemd; dit zijn organismen met genetisch verschillende weefsels (naar het bekende Griekse mythologische wezen; een fusie tussen verschillende dieren).De chimeren bestonden uit 2, 3 of zelfs 6 verschillende kolonies (figuur 1). Chimeren bestaande uit 4 of meer genetisch verschillende kolonies werden multi-partner entities (meerdere-partners entiteit, of MPE) genoemd. Verder zag men 2 of 3 kolonies welke elkaar tegelijk afstootten. Na fusie was bijna niet meer te zien wat de oorspronkelijke kolonies waren, hoewel niet alle fusies permanent waren. De afstotingsverschijnselen waren divers; afwezigheid van weefselverbinding waarbij skeletten elkaar raakten, overgroei van één van de kolonies, bleking, necrose (afsterving) en verder dode kolonies. Aanvankelijk kwamen fusie- en afstotingsverschijnselen even vaak voor, maar naarmate de chimeren uit meer partners bestonden nam dit laatste af. Hoe groter de chimeer, hoe kleiner de kans dat een nieuwe kolonie werd afgestoten.
Figuur 1: Stylophora pistillata kolonies van 0 tot 4 maanden oud, op zeer korte afstand naast elkaar geplaatst, vormden chimeren of stootten elkaar af. a: enkel genotype, 2 maanden oud. b: bi-chimeer; 2 gefuseerde kolonies c: 2 genotypen welke elkaar afstootten, d:tri-chimeer, 1,5 maand oud, e: 3 elkaar afstotende genotypen, f: samengestelde kolonie van 7 genotypen; een zogenaamde multi-partner entity (meerdere-partners entity of MPE). Opvallend is dat slechts 1 genotype wordt afgestoten, geheel links. Asteriksen geven afstoting aan, stippellijnen geven grenzen tussen oorspronkelijke kolonies aan, pijlen geven de primaire poliepen (poliepen waaruit de kolonie is ontstaan, na metamorfose) aan. De witte schaalbalken rechts onderaan zijn 1 mm (Shaish et al, PloS ONE, 2008).Deze resultaten zijn heel interessant, aangezien de wetenschappers eerdere onderzoeken naar immuniteit bij koralen hiermee bevestigden. Koralen hebben duidelijk een subtiele manier gevonden om vreemd van eigen weefsel te onderscheiden; een sterk kenmerk van een immuunsysteem. Verder kan worden geconcludeerd dat deze immuniteit is aangeboren, zoals bij veel dieren het geval is. Dit is zo omdat deze koralen nog niet eerder in contact waren gekomen met vreemd weefsel. In dit geval noemt men deze reacties op vreemd weefsel een allogene reactie; dit betekent dat het koraal reageert op genetisch verschillend weefsel van dezelfde soort. Het tegengestelde is isogeen; genetisch identiek, zoals een transplantatie van een orgaan tussen een één-eiige tweeling, en autogeen; zoals een transplantatie van het eigen weefsel (bijvoorbeeld huid) naar een andere locatie.
Zodra planula larven van deze soort zich op het rif vestigen, dan vindt in feite bovenstaand experiment plaats. De koralen reageren op elkaars weefsel, en dit zal zowel intra- als interspecifiek plaatsvinden (interacties tussen de eigen soort en andere soorten). Uiteindelijk zullen zich zowel solitaire kolonies als chimeren gaan vormen, hoewel het laatste een zeldzamer geval lijkt. Verder werd gevonden dat de gemiddelde grootte van de MPE- (4 of meer gefuseerde kolonies) leden kleiner was dan die van de solitaire kolonies (hoewel de MPE’s in totaal uiteraard wel groter waren). Dit is waarschijnlijk het resultaat van de interacties tussen de kolonies, zoals afstoting. Dit verbruikt veel energie, wat ten koste gaat van de groei. Het feit dat de MPE’s groter waren is een indicatie voor verhoogde overlevingskansen. De les die hieruit kan worden getrokken is dat hoewel een verbond tussen organismen voordelen heeft, er hiernaast ook nadelen aan kleven. Als er inderdaad kosten zijn zoals groei-inhibitie (remming) en zelfs sterfte, waarom vormen koralen dan toch deze MPE’s, of “superkolonies”? Overleven op het rif draait allemaal om het innemen van ruimte, en bij MPE’s werd gevonden dat deze sneller groeiden. Een snellere groei betekent het sneller innemen van een niche, een specifieke plaats, op het rif. Dit verhoogt vervolgens de overlevingskansen van deze kolonies, omdat zij binnen korte tijd een plek veroverd hebben waar zij licht kunnen opvangen en nutriënten kunnen opnemen.
Deze resultaten laten zien dat koralen absoluut bijzondere dieren zijn. Regelmatig worden nieuwe manieren gevonden waarop koraalpoliepen samenwerken. Niet alleen kloneren (delen) veel soorten zich waarbij ze grote kolonies vormen, ze fuseren ook met genetisch verschillende individuen om hun overlevingskansen te vergroten. Dit verschijnsel, natuurlijk chimerisme, is ook waargenomen bij veel andere sessiele (vastzittende) mariene organismen. Voorbeelden hiervan zijn sponzen, zachte koralen, zakpijpen en zelfs bepaalde steenkoralen.Lobophyllia corymbosa
Naast Stylophora pistillata, is ook bij andere koraalsoorten immuniteit ontdekt, waaronder bij de steenkoraalsoort Lobophyllia corymbosa2. Wetenschappers van de Universiteit van Tel Aviv en de nationale Universiteit van Ierland ontdekten recentelijk dat ook poliepen van L. corymbosa vreemd van eigen weefsel kunnen onderscheiden. Verder zagen zij ook dat deze poliepen nutriënten transporteerden naar verwonde exemplaren binnen de eigen kolonie.
De soort L. corymbosa lijkt wat op soorten van het genus Caulastrea; wanneer de kolonie volgroeid is, snoeren de poliepen hun weefsel volledig af waardoor deze solitair worden. De onderzoekers verwijderden 52 poliepen van 14 verschillende kolonies te Eilat, Israël. De poliepen werden of intact gelaten, of in de lengte vanaf boven doorgesneden. Vervolgens werden de poliepen, of helften daarvan, op verschillende manieren aan elkaar verbonden met koperdraad. Dit leverde 3 vormen van transplantatie op; autotransplantaties (twee poliephelften van de originele poliep), isotransplantaties (twee poliephelften van verschillende poliepen, maar van dezelfde kolonie, welke dus genetisch identiek zijn) en allotransplantaties (twee helften van genetisch verschillende poliepen).
Alle auto- en isotransplantaties vertoonden complete fusie van zowel koraalweefsel als skelet, binnen 6 weken! Dit werd bevestigd door histologische coupes van gefuseerde poliepen. Dit zijn poliepen die zijn ingebed in paraffine (een was), zeer dun zijn gesneden en vervolgens zijn gekleurd en bekeken onder de microscoop. Daarentegen vond er nergens fusie plaats tussen de genetisch verschillende allotransplantaties. Er was of geen reactie van de helften, waarbij de veroorzaakte wonden zich heelden, of zij stootten elkaar heftig af. De manier van afstoting is volgens de onderzoekers nog niet geheel duidelijk. Dit is waarschijnlijk een cytotoxische afweer, waarbij beide helften elkaar belagen met giftige stoffen (denk aan steroïden, terpenen en andere organische stoffen). Ook netelcellen kunnen mede verantwoordelijk zijn.
 Deze resultaten zijn volledig in samenspraak met die van Stylophora. Deze immuniteit is ook gevonden bij Fungia scutaria3. Andere wetenschappers voerden experimenten met F. scutaria uit, die lieten zien dat ook zij lichaamsvreemd weefsel afstootten. Twee helften van dit koraal fuseerden slechts wanneer deze van anthocauli (klonen die geproduceerd worden wanneer een Fungia poliep afsterft) afkomstig waren. Een transplantatie van twee genetisch verschillende poliepen leidde altijd tot wederzijdse afstoting.
Deze resultaten zijn volledig in samenspraak met die van Stylophora. Deze immuniteit is ook gevonden bij Fungia scutaria3. Andere wetenschappers voerden experimenten met F. scutaria uit, die lieten zien dat ook zij lichaamsvreemd weefsel afstootten. Twee helften van dit koraal fuseerden slechts wanneer deze van anthocauli (klonen die geproduceerd worden wanneer een Fungia poliep afsterft) afkomstig waren. Een transplantatie van twee genetisch verschillende poliepen leidde altijd tot wederzijdse afstoting. Het feit dat poliepfusie alleen plaats vindt na weefselschade (zoals bij het halveren van poliepen) is te verklaren doordat de regeneratieve processen in het koraalweefsel worden ingeschakeld. Koralen bezitten namelijk stamcellen, welke actief worden na weefselschade. Wanneer deze delen, ontstaan nieuwe cellen; bindweefsel-, spier-, zenuw- en huidcellen. Uiteindelijk leidt dit tot de opbouw van nieuw weefsel, en de complete fusie van genetisch identiek koraal. Het is heel intrigerend waarom koralen en andere lagere dieren dit vermogen hebben, terwijl hogere gewervelde dieren dit in veel mindere mate bezitten. Vanuit een evolutionair perspectief lijkt dit onvoordelig te zijn.
Energietransport
Een volgende stap in het onderzoek was het bepalen van de mate van energietransport binnen een enkele L. corymbosa kolonie. Hierbij werd gekeken naar het transport van koolhydraten, afkomstig van fotosynthese. Het is bekend dat veel koralen nutriënten transporteren binnen de kolonie, via het gemeenschappelijke weefsel (coenenchym of coenosarc genoemd). Dit komt onder andere voor bij Montastrea en Porites soorten. Om dit te bepalen werd een zeer creatief experiment opgezet; poliepen werden verwijderd en geïncubeerd in water met radioactief koolstof (14C), waarbij zij 20 uur per dag werden belicht. Hierdoor bouwden de koralen veel 14C in hun weefsels in, via een hoge mate van fotosynthese. Hierna werden deze poliepen teruggeplaatst in de eigen kolonie (isogeen koraal), of een genetisch verschillende (allogeen koraal). Van elke kolonie werd één poliep beschadigd (verwonding van 2 cm2). Een aantal poliepen werd ook in plastic gewikkeld om het effect hiervan te bepalen. Het experiment is in de afbeelding hieronder weergegeven:
Schematische weergave van het 14C labeling experiment van L. corymbosa poliepen. 48 uur na de terugplaatsing van de zogenaamde ‘hot polyp’ (radioactieve poliep) werden 4 fragmenten verwijderd (weergegeven door de zwarte cirkels) met een stalen boor. Hiermee werden weefselfragmenten van 1 cm2 elk verwijderd. Één fragment werd genomen van de ractioactieve poliep, één van de verwonde poliep, één van een intacte poliep dichtbij de radioactieve poliep, en één hier verder vanaf. De radioactiviteit van de monsters werd gemeten met een vloeistof scintillatieteller (Bricker et al, Journal of Experimental Biology, 2006).
De onderzoekers vonden dat L. corymbosa poliepen alleen nutriënten (in elk geval koolhydraten) transporteerden naar genetisch identieke, verwonde poliepen. Er werd geen radioactiviteit gevonden bij isogene intacte, of allogene poliepen (zowel intact als verwond). Dit is opmerkelijk, en past heel goed bij de eerder besproken resultaten. Koralen kunnen niet alleen genetisch identieke poliepen herkennen, zij helpen deze ook na verwonding door het transporteren van nutriënten naar een locatie met extra vraag naar energie. Nutriënten zoals koolhydraten kunnen het herstel van verwonde poliepen versnellen. Het is niet verrassend dat poliepen slechts elkaars klonen helpen, omdat zij alleen hier een evolutionair voordeel bij hebben. Een gehele koraalkolonie is genetisch geheel identiek, en alle leden van deze kolonie hebben als doel het verspreiden van hun genen naar volgende generaties. Het helpen van een naburige poliep is feitelijk het helpen van de poliep zelf.
Een volgende vraag was hoe het mogelijk was dat koolhydraten getransporteerd konden worden tussen poliepen. Dit lijkt voor de hand liggend, maar L. corymbosa poliepen zijn niet met elkaar verbonden door weefsel. De poliepen kunnen dus eigenlijk als solitair worden beschouwd, hoewel dit experiment liet zien dat zij zich toch als één kolonie gedroegen. Deze poliepen bepaalden zelfs de richting van het nutriëntentransport. Hoe is dit mogelijk?
De wetenschappers hebben twee mogelijke verklaringen voor dit verschijnsel. De eerste is dat koraalmucus (slijm) door de polieptentakels wordt verplaatst, in een specifieke richting. De tweede verklaring is dat loslatende cellen van de donorpoliep zich verplaatsen (of worden verplaatst) naar de ontvangende. Een combinatie van deze is uiteraard ook mogelijk. Het is namelijk zo dat deze poliepen zich ’s nachts uitstrekken waardoor zij elkaar raken. Tijdens deze nachtelijke periode zou het transport kunnen plaatsvinden. Deze theorieën worden bevestigd door het feit dat in plastic gewikkelde poliepen geen energietransport laten zien, ook niet naar verwonde poliepen.
Bovenstaande resultaten laten zien dat L. corymbosa poliepen sterk geïntegreerd zijn; zij recyclen energie naar poliepen welke dit nodig hebben, en onderscheiden vreemde van genetisch identieke poliepen. De vraag is waarom sommige koraalsoorten kiezen voor een puur koloniale levensstijl (zoals de genera Acropora, Montipora en Stylophora), waar andere soorten juist solitair blijven (denk aan Trachyphyllia, Lobophyllia en Caulastrea). Volgens de onderzoekers hebben beide vormen hun voordelen. Kolonievorming is ideaal voor het transport van energie tussen poliepen; L. corymbosa poliepen waren niet in staat energie over een langere afstand te verspreiden, in tegenstelling tot koloniale soorten. Echter, een solitaire vorm biedt weer meer bescherming tegen gevaarlijke infecties zoals door Vibrio bacteriën (denk aan een Montipora die geheel afsterft , wat deels kan worden voorkomen door fragmentatie). Recent is gevonden dat deze schadelijke bacteriën verschillende vormen van ‘white band syndrome’ veroorzaken, wat leidt tot bleking en RTN (rapid tissue necrosis; het zeer snel afsterven van koraalweefsel)4. Een tweede voordeel van een solitaire levensstijl is dat het verspreiden van genetisch materiaal sneller verloopt. L. corymbosa poliepen breken bijvoorbeeld makkelijker af waardoor zij sneller nieuwe plaatsen op het rif kunnen koloniseren. Fungia poliepen bewegen zich langzaam voort, en worden tevens door stroming verspreid. Uiteindelijk draait het in de biologie allemaal om overleving en voortplanting...
Ter conclusie
Aquaria thuis worden vaak vol geplaatst met vele soorten koraal, op kleine afstand. Wanneer deze koralen elkaar gaan overgroeien en netelen levert dit nogal eens gebleekte en dode koralen op. Als u dit thuis ziet, denkt u dan nog eens aan dit verhaal, en aan de bijzondere processen die zich hierbij afspelen.
Referenties
Keren-Or Amar, Nanette E Chadwick and Baruch Rinkevich, Coral kin aggregations exhibit mixed allogeneic reactions and enhanced fitness during early ontogeny, BMC Evolutionary Biology, april 2008, 30;8:126
Itzchak Brickner, Uri Oren, Uri Frank and Yossi Loya, Energy integration between the solitary polyps of the clonal coral Lobophyllia corymbosa, Journal of Experimental Biology, 2006, 209, pp 1690-1695
Jokiel PL Bigger CH, Aspects of histocompatibility and regeneration in the solitary reef coral Fungia scutaria, Biol. Bull., 1994, 186(1):72-80
Meir Sussman, Bette L. Willis, Steven Victor, David G. Bourne, Coral Pathogens Identified for White Syndrome (WS) Epizootics in the Indo-Pacific, PloS ONE, 2008, 3(6)
Reactie van steenkoralen op licht van verschillende golflengte en sterkte.
Dit artikel geeft een indruk hoe en waarom bepaalde koralen reageren (of niet) op lichtsterkte en de golflengte van dit licht. Dit is van belang om te begrijpen welke soort licht we de koralen moeten geven om ze maximaal te voeden, zodat ze ook in onze aquaria gezond blijven. Het laat ook zien waarom het kleurenspectrum van de lampen boven het aquarium zo belangrijk is.
O. Levy*, Z. Dubinsky and Y. Achituv: Photobehavior of stony corals: responses to light spectra and intensity, The Journal of Experimental Biology 206, 4041-4049 (2003)
Vertaling: Rien van Zwienen
Inleiding
 Binnen de steenkoralen, gorgonen en anemonen is er verschillend gedrag wat betreft het samentrekken en uitzetten van de poliepen en tentakels. De meeste steenkoralen expanderen hun poliepen alleen ’s nachts. Slechts een paar soorten expanderen hun poliepen gedurende de dag en enkelen hebben hun poliepen altijd uit staan. Koralen die hun poliepen ’s nachts openen, houden die tot de schemering open. Echter, een lichtbundel of aanraking kan er voor zorgen dat de tentakels onmiddellijk weer intrekken.
Binnen de steenkoralen, gorgonen en anemonen is er verschillend gedrag wat betreft het samentrekken en uitzetten van de poliepen en tentakels. De meeste steenkoralen expanderen hun poliepen alleen ’s nachts. Slechts een paar soorten expanderen hun poliepen gedurende de dag en enkelen hebben hun poliepen altijd uit staan. Koralen die hun poliepen ’s nachts openen, houden die tot de schemering open. Echter, een lichtbundel of aanraking kan er voor zorgen dat de tentakels onmiddellijk weer intrekken.
Zowel steenkoralen als diverse andere rif organismen bevatten eencellige algen, genaamd zoöxanthellen, die in een groot gedeelte van hun energiebehoefte voorzien via fotosynthese. Koralen zijn vleeseters en omdat zoöplankton vooral ’s nachts aanwezig is, wordt er aangenomen dat de meeste koralen hun tentakels expanderen om dan hun prooien te vangen. De mate van expansie zou beïnvloed kunnen worden door stroming en aanwezigheid van voedsel.
In anemonen met zoöxanthellen is er verband tussen fotosynthese en de mate van expansie. Anemonen met veel zoöxanthellen zijn voortdurend uitgezet, terwijl dieren met weinig of geen zoöxanthellen gedurende de dag instaan.
Er is onderzoek gedaan naar het expansie gedrag van het Montastrea cavernosa koraal . In ondiep water blijven veel poliepen gedeeltelijk uit staan, dit wordt wel dag-gedrag genoemd. Nachtgedrag of tentakel expansie gedurende de nacht werd op dieptes groter dan 20 meter gezien. Op die dieptes werd er zelden overdag poliep expansie gezien. Dit verschillende gedrag is waarschijnlijk te verklaren door het verschil in zoöxanthellen dichtheid. Er wordt aangenomen dat dagactieve kolonies meer zoöxanthellen hebben dan de nachtactieve.
Koralen kunnen goed gedijen in voedselarme wateren vanwege hun symbiose met zoöxanthellen. Aan hun koolstof en energie behoeftes wordt voldaan met verschillende soort- specifieke fotosynthetische producten en door het vangen van zooplankton, in sommige gevallen aangevuld met kleine bijdragen van opgeloste organische stoffen en bacteriën.
Op het koraalrif bij Eilat, in de noordelijke Rode Zee, zetten de Favia favus en Plerogyra sinuosa koralen ’s nachts hun poliepen uit en trekken die weer in bij de dageraad.
Goniopora lobata en Stylophora pistillata hebben hun poliepen altijd uit staan.
 |
 |
 |
 |
 |
| Favia favus | Goniopora lobata | Plerogyra sinuosa | Cladopsammia gracilis | Stylophora pistillata |
Fig.1 vijf onderzochte koraalsoorten
-geel.jpg) De licht omgeving is een belangrijke factor bij de productiviteit, fysiologie en ecologie van koralen. Onderwater neemt het licht exponentieel af met de diepte, volgt ruwweg de wet van Lambert-Beer. Het licht wordt onderwater ook verzwakt door opgeloste en zwevende deeltjes, en heel belangrijk, door fytoplankton. Licht verzwakking is niet gelijk voor alle golflengtes, en de waterkolom werkt als een monochromator, verkleint het spectrum van het verst doordringende licht tot een relatieve nauwe bandbreedte. In de heldere wateren rond het tropisch rif, is de uitdoving van het violet en blauwe gedeelte van het spectrum minimaal, terwijl de uitdoving groter is bij langere golflengtes (rood). Echter, in zulke “blauwe woestijn achtige” ondiepe wateren kunnen koralen ook blootgesteld worden aan behoorlijke hoeveelheden rode golflengtes en het niet zichtbare UV licht.
De licht omgeving is een belangrijke factor bij de productiviteit, fysiologie en ecologie van koralen. Onderwater neemt het licht exponentieel af met de diepte, volgt ruwweg de wet van Lambert-Beer. Het licht wordt onderwater ook verzwakt door opgeloste en zwevende deeltjes, en heel belangrijk, door fytoplankton. Licht verzwakking is niet gelijk voor alle golflengtes, en de waterkolom werkt als een monochromator, verkleint het spectrum van het verst doordringende licht tot een relatieve nauwe bandbreedte. In de heldere wateren rond het tropisch rif, is de uitdoving van het violet en blauwe gedeelte van het spectrum minimaal, terwijl de uitdoving groter is bij langere golflengtes (rood). Echter, in zulke “blauwe woestijn achtige” ondiepe wateren kunnen koralen ook blootgesteld worden aan behoorlijke hoeveelheden rode golflengtes en het niet zichtbare UV licht.
Recentelijk is aangetoond dat koralen met zoöxanthellen, zelfs blauw maanlicht, dat bestaat uit de verst doordringende golflengtes in gebieden waar koraalriffen voorkomen, opnemen.
Het doel van deze studie was om uit te vinden of het expanderen en intrekken van poliepen van koralen met zoöxanthellen gebeurt als een directe reactie op licht, of als een indirecte reactie, geïnitieerd door de fotosynthetische activiteit van de symbiotische algen.
We onderzochten de mogelijkheid dat de expansie en intrekking van tentakels de fotosynthese optimaliseert.
We onderzochten expansie en intrekken als gevolg van verschillende licht intensiteiten en golflengtes over verschillende periodes, inclusief het geen zoöxanthellen bevattende koraal Cladopsammia gracilis.
We bestudeerden de absorptie en actie spectra voor fotosynthese en verdeling van zoöxanthellen binnen de koralen. Tevens werden de fotosynthetische karakteristieken van de vier bestudeerde koralen bestudeerd.
Stukken koraal van de volgende rifbouwende steenkoralen: Favia favus, Goniopora lobata, Stylophora pistillata en Plerogyra sinuosa en het geen zoöxanthellen bevattende koraal Cladopsammia gracilis werden van 5-7 meter diep van het koraalrif bij Eilat gehaald. Deze kolonies (kleiner dan 6 cm) werden in het laboratorium in een ondiep aquarium geplaatst. Na een gewenningsperiode vertoonden ze hetzelfde expansie/intrek gedrag als hun niet verplaatste broertjes/zusjes op het rif.
Resultaten
Het meeste licht, op
5 meter diepte, op een heldere dag, is rond het middaguur, 12-13 uur (zie fig. 2. Er is een kleine afname bij golflengtes > 600 nm. Er was toch een behoorlijke penetratie van alle golflengtes tussen 400 en 700 nm bij 5 meter diepte. De verdeling van het licht onderwater, varieert gedurende de dag door veranderingen van de invalshoek van het zonlicht en rond het middaguur dringt er bij alle golflengtes het meeste licht door.
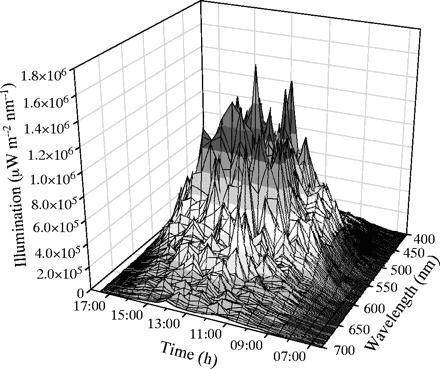
Fig 2. 3-dimensionaal beeld van licht opbrengst op 5 meter
diepte in de golf van Eilat.
|
violet |
400 tot 450 nm |
|
blauw |
450 tot 500 nm |
|
groen |
500 tot 550 nm |
|
geel |
550 tot 600 nm |
|
oranje |
600 tot 650 nm |
|
rood |
650 tot 700 nm |
Tabel 2 Golflengtes (nm) van verschillende kleuren licht
Poliepen van de Favia favus trekken zich binnen 5-6 minuten bij lage lichtintensiteiten volledig in bij golflengtes van 400-520 nm. (fig.3A,B)
Bij hoge lichtintensiteiten reageren de poliepen bij golflengtes tussen 660 en 700 nm. Poliep contractie tussen 540 en 640 nm was erg langzaam onafhankelijk van de hoeveelheid licht.
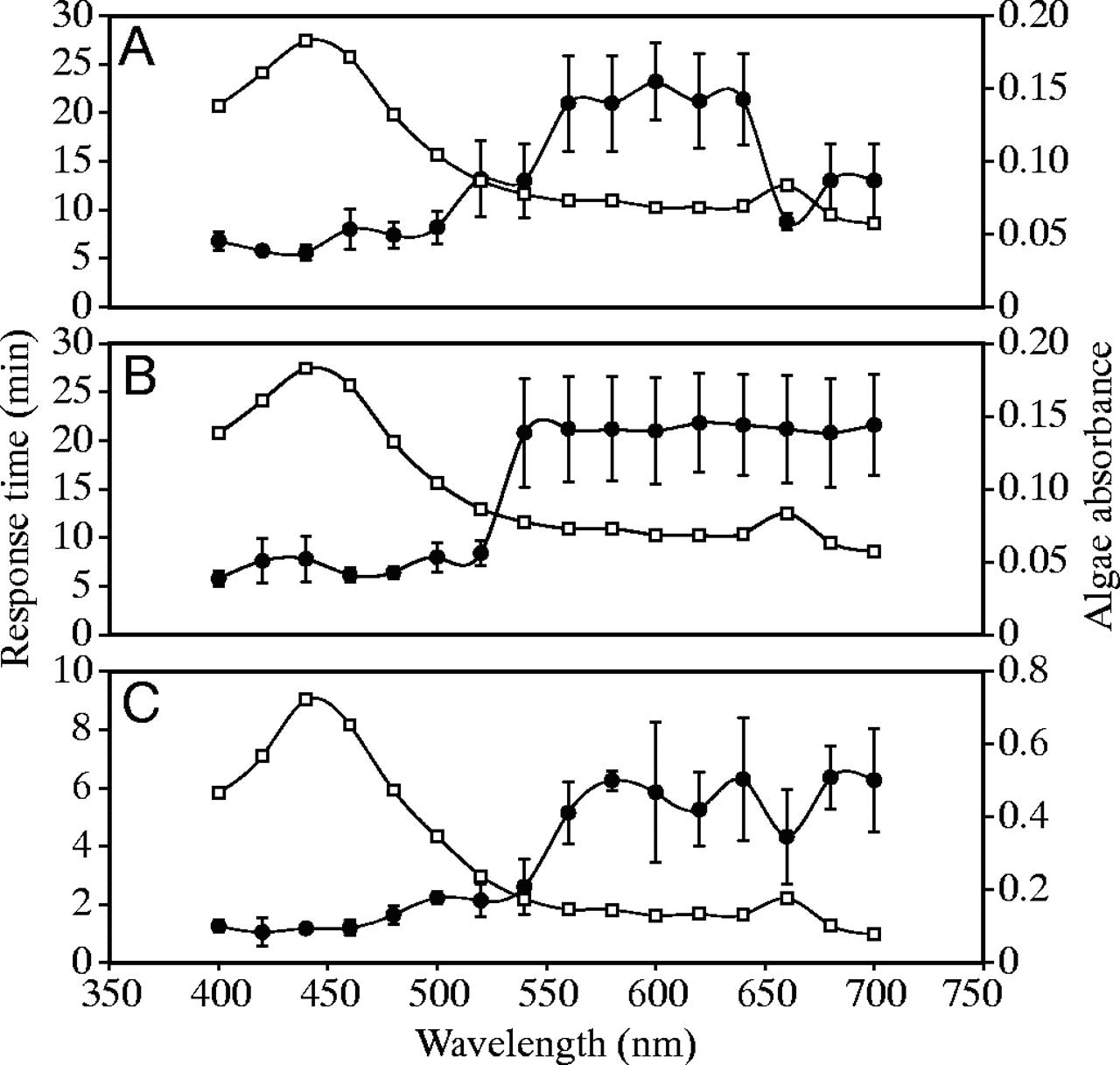
Fig. 3. Actie diagram voor poliep contractie bij Favia favus and Plerogyra sinuosa (zwarte rondjes). (A) vergelijking F. favus poliep contractie tegen lichtspectrum geabsorbeerd bij zoöxanthellen bij grote lichtsterkte (openvierkantjes) (B) vergelijking F. favus poliep contractie tegen lichtspectrum geabsorbeerd bij zoöxanthellen bij lage lichtsterkte (openvierkantjes).(C) vergelijking P. sinuosa poliep contractie tegen lichtspectrum geabsorbeerd bij zooxanthellen bij grote lichtsterkte (openvierkantjes).
Goniopora lobata, Stylophora pistillata en Cladopsammia gracilis reageerden helemaal niet op licht, van welke golflengte dan ook.
Kolonies van Plerogyra sinuosa, blootgesteld aan veel licht, trekken hun poliepen in bij golflengtes tussen 400-540 nm. Dit koraal reageerde ook op licht van 660nm (fig. 3C). Golflengte van 400-540 nm had een duidelijk anders effect dan 560-700 nm.
Bij koralen die reageerden op de lichtprikkels, werden de golflengtes waarbij ze het meest effectief waren om hun poliepen in te trekken in verband gebracht met de absorptie spectra van hun symbiotische algen (fig. 3A en B) Bij P.sinuosa, was het verband tussen de spectrale absorptie van de zoöxanthellen en het actie spectrum van de poliepen significant.
Poliep contractie in het koraal Cladopsammia gracilis (zonder zoöxanthellen) gebeurde niet onder invloed van licht, van welke golflengte dan ook. Zelfs zeer grote lichtsterktes hadden geen effect.
De spectrale gegevens van de verschillende pieken laten zien dat meeste van de belangrijkste pigmenten behoorlijke absorptie hebben tussen 400-540 nm, met grootste pieken tussen 440-480 nm. Het breedste absorptie spectrum profiel behoort tot de carotenoïde pigmenten, zoals perdinin, diatoxanthin en diadinoxanthin, die blauw/blauwgroene absorptie banden laten zien die gedeeltelijk de chlorofyl banden overlappen (figuur 4).
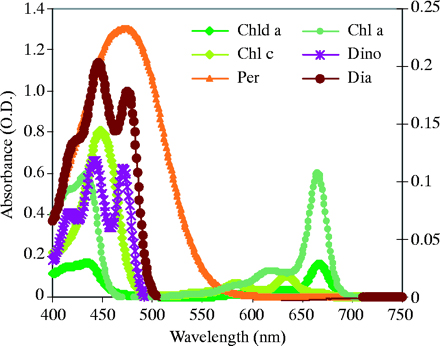
Figuur 4. Absorptie karakteristiek van de belangrijkste pigmenten geëxtraheerd van F. favus zoöxanthellen. Pigmenten zijn: Chl, chlorophyll; Chld, chlorophyllidae; Per, perdinin; Dia, diatoxanthin; Dino, diadinoxanthin
ATI T5 54W 12000K Aquablue special ATI T5 39W Blue Plus
Kleurenspectrum van twee ATI T5 lampen
De hoogste dichtheid aan zoöxanthellen werd gevonden in de poliepen van Goniopora lobata (1.78±0.58x106 cellen per cm2). Lagere dichtheden werden gevonden in de poliepen van Plerogyra sinuosa. In de poliepen van Favia favus of Stylophora pistillata werden geen zoöxanthellen gevonden (tabel 1), alhoewel er soms zoöxanthellen werden gezien als de poliepen onder de microscoop onderzocht worden.
Coral Species Number of colonies Cells in total coral tissue (cells x 106/cm2) Cells in the tentacles (cells x 106/cm2) Cells in tentacles/Cells total
Stylophora. pistillata 4 0.93 ±0.07 0 0
Favia favus 5 0.34 ±0.19 0 0
Plerogyra sinuosa 4 1.96 ±1.11 0.47 ±0.05 0.238
Goniopora lobata 4 3.25 ±0.93 1.78 ±0.58 0.547
Metingen aan de fotosynthese activiteit laten zien dat de maximale chlorofyl fluorescentie verschillen het grootst zijn in het blauwe gebied. In alle vier de soorten werden de laagste
waarden gemeten als de koralen werden belicht met golflengtes van 540-620nm (figuur 5)

Foto: Bas Arentz
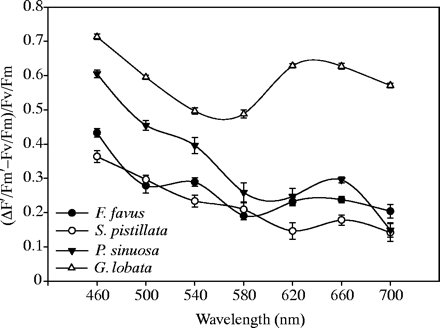
Figuur 5. Effect van het licht spectrum (460-700nm) op de opbrengst van de fotosynthese bij vier koraal soorten: Favia favus; Stylophora pistillata; Plerogyra sinuosa; Goniopora lobata.
Discussie
De reactie van Favia favus en Plerogyra sinuosa op licht van verschillende golflengte hangt af van het absorptie spectrum van hun symbiotische zoöxanthellen. Er zijn voldoende aanwijzingen dat actie spectra van fotosynthetische pigmenten in de buurt liggen van die van hun absorptie spectra. De afname van de opbrengst in verschillende fotosynthese actie
spectra werd ook in verband gebracht met het intrekken van poliepen. Dit verband kan er op duiden dat het intrekken van poliepen wordt veroorzaakt, minstens indirect, door de
fotosynthese van de zoöxanthellen. Dit verband komt overeen met de observaties van Lasker, die zag dat belichte kolonies van Montastrea cavernosa
niet meer uit gingen staan als hun zoöxanthellen gebleekt of weg waren. Ook Sawyer liet zien dat bij anemonen de reactie op licht bij soorten die zoöxanthellen bevatten intenser is dan bij soorten die geen zoöxanthellen bevatten.

Foto: Mark Breugelmans
In de poliepen van Favia favus en Stylopora pistillata werden bijna geen algen waargenomen, en vandaar de poging hun dichtheid te bepalen faalde (tabel 1). We concludeerden dus dat het aantal zoöxanthellen in Favia favus en Stylopora pistillata verwaarloosbaar is. Daarom zal de vorm van hun poliepen hun fotosynthese snelheid nauwelijks beïnvloeden, wat bevestigd werd door de metingen. Daarentegen, werden de grote hoeveelheid zoöxanthellen in de poliepen van Plerogyra sinuosa gemakkelijk gemeten (tabel 1). De dichtheid van algen cellen in de poliepen in verhouding tot de totale gemiddelde dichtheid in het koraal weefsel was veel hoger bij G. lobata dan in de drie andere onderzochte koralen.
De twee soorten met lage zoöxanthellen dichtheid (F. favus en P. sinuosa) bleven samengetrokken gedurende de dag, terwijl de koralen met hoge zoöxanthellen dichtheid hun poliepen expandeerden als ze belicht werden. De verschillende poliepstructuur van P. sinuosa en F. favus ten opzichte van G. lobata houdt in dat hun poliepen ook verschillende taken hebben. De vingervormige poliepen van de eerste twee hebben als doel prooi te vangen. Ze hebben waarschijnlijk weinig of geen betekenis in de controle van het beschikbare licht wat aanwezig is voor fotosynthese in de zoöxanthellen en expanderen alleen ’s nachts. Daarentegen, expanderen de bloemvormige tentakels van G. lobata gedurende de dag wel om licht te absorberen, wat ook te zien is aan de zoöxanthellendichtheid in het weefsel van hun tentakels. Als de tentakels van G. lobata open staan, wordt het oppervlak van de kolonie ongeveer 7.5 keer zo groot. Zo’n grote toename van het oppervlak gedurende de expansie ondersteund de hypothese dat koralen met een hoge zoöxanthellen dichtheid graag open willen staan gedurende de dag om efficiënt licht te absorberen, wat er voor zorgt dat er meer energie naar het koraal gaat.

Foto: Hans Peter

Foto: Reefsecrets
P. sinuosa kolonies bestaan uit voedsel tentakels en bolvormige blaasjes. De blaasjes bevatten grote aantallen zoöxanthellen, maar bevatten geen netelcellen. De bolvormige blaasje expanderen overdag, terwijl de tentakels die netelcellen bevatten ’s nachts expanderen, en de blaasjes worden groter met toenemende diepte, om zodoende te compenseren voor de afname van het licht. De afwezigheid van netelcellen in de blaasjes met hoge zoöxanthellen dichtheid en het feit dat tentakels die netelcellen bevatten slechts weinig zoöxanthellen bevatten, wijzen erop dat de rol van de bolletjes is om licht te vangen, zoals bij tentakels van G. lobata, terwijl de tentakels van P. sinuosa alleen dienen om voedsel te vangen.
Expansie van tentakels met weinig zoöxanthellen kan leiden tot netto energie verlies, omdat expansie energie kost. Tevens kan expansie van tentakels met weinig zoöxanthellen leiden tot afname van de fotosynthese snelheid door licht verstrooiing.
Sommige zeeanemonen die zoöxanthellen bevatten hebben twee typen gespecialiseerde organen: pseudo-tentakels, met hoge concentratie zoöxanthellen, en echte tentakels met weinig of geen algen. De pseudotentakels expanderen overdag en zijn fotosynthetisch actief. De echte tentakels expanderen ’s nachts en worden gebruikt om zooplankton te vangen. Het koraal Montastrea cavernosa, dat twee verschijningsvormen heeft, vertoont vergelijkbaar gedrag. Kolonies met een dichte zoöxanthellen dichtheid hebben de neiging overdag open te staan, terwijl de vorm met weinig zoöxanthellen alleen ’s nachts open staat.
We denken dat verschillen in algen dichtheid en hun verdeling in het weefsel kan leiden tot verschillen in bijdrage aan hun energiebronnen.
Onze resultaten op het effect van licht spectrum op samentrekking van tentakels bij F. favus and P. sinuosa en de afwezigheid van dit effect bij koralen die geen zoöxanthellen hebben, steunen de gedachte dat dit gedrag te maken heeft met de fotosynthetische activiteit van de zoöxanthellen.

Foto: Luc Loyen

Foto: Germain Leys
Metingen laten zien dat de twee soorten die overdag in staan een behoorlijk lagere fotosynthetische activiteit hebben als hun tentakels uitgezet zijn dan wanneer zij zijn ingetrokken. Bij G. lobata en S. pistillata is er, onafhankelijk van hun vorm van de tentakels, geen verschil in fotosynthetische activiteit. Men neemt aan dat de expansie-samentrekking van de poliepen als een soort diafragma werkt, die de fotosynthese van de koraal kolonie optimaliseert. Maximale fotosynthetische respons is in het blauwgroene gebied, dat overeenkomt met de maximale lichtdoorlatendheid van tropische wateren. Ons onderzoek lijkt erop te wijzen dat er een verband is tussen fotosynthese van de zoöxanthellen (of de producten hiervan) en gedrag van tentakels in koralen, maar kunnen niet uitsluiten dat het mogelijk is dat zulke cellen gevoelig zijn voor dezelfde golflengten als de fotosynthese, en dat dit een rol speelt in het gedrag van de koralen.
Inloggen
Registreren
Foto van de maand

Blauwe mandarijnpitvis Synchiropus splendidus
Foto: Sander Hemmes