Portaalsite voor de échte zeeaquariaan
Zoeken op de site
Sponsors







Partners








Tweeslachtigheid van vissen
Tweeslachtigheid van zeevissen
Tim Wijgerde (vertaling Germain Leys)
Wist u dat veel zeevissoorten van geslacht kunnen veranderen als dat nodig is? Wetenschappers hebben ontdekt dat de soorten Pseudochromis flavivertex, Pseudochromis aldabraensis en Pseudochromis cyanotaenia van geslacht kunnen veranderen in een éénslachtige omgeving.
Dit betekent in feite dat wanneer twee vrouwtjes bij elkaar worden geplaatst, één ervan een mannetje wordt (of andersom). Als een individu verandert van man naar vrouw, noemen we dit proterandrie. Als het verandert van vrouw naar man, noemen we het protogynie

Pseudochromis aldabraensis Foto Germain Leys
Dieren die zowel mannelijke (testes) als vrouwelijke (eierstokken) geslachtsorganen hebben heten hermafrodieten. Deze vissen zijn zogenaamde sequentiële hermafrodieten, net zoals de anemoon vissen van de genera Amphirion en frenatus, omdat ze niet zowel mannelijk als vrouwelijk tegelijk zijn.
De aquariumkweek van zeevissen profiteert van dit geslachtsveranderend vermogen, waardoor het minder moeilijk is om een broedpaar te verkrijgen. Helaas, de pas uitgekomen jongen dienen gevoederd te worden met zoöplankton-culturen om te overleven.

Pseudochromis flavivertex Foto: Germain Leys
Een ander mooi voorbeeld van sequentiële hermafrodisme wordt weergegeven door de dwergkeizers (Centropyge spp.). Ze veranderen van geslacht van vrouw naar man als het dominante mannetje van de groep verdwijnt of overlijdt, dus ze zijn protogynische soorten. Wanneer een man van een harem verdwijnt of overlijdt, dan zal het grootste vrouwelijke exemplaar van geslacht veranderen binnen een paar weken!
Onlangs bleek dat Centropyge ferrugata mannen terug keerden naar het vrouwelijke stadium nadat ze gedomineerd werden door een groter en dominanter mannetje. Ze veanderden terug in hun vrouwelijke staat binnen 47-89 dagen. Niet alleen hebben ze hun geslacht veranderd, hun uiterlijk werd ook veranderd. Keizervissen vertonen wat seksueel dichromatisme wordt genoemd, of seksueel dimorfisme. Dit betekent dat mannen en vrouwen er anders uitzien (de mens is een heel mooi voorbeeld). Mannelijke dwergkeizers Centropyge ferrugata hebben een specifieke kleuring van de rugvin. Nadat de man naar vrouw overgang, was zelfs de rugvin veranderd naar die van een vrouw.

Centropyge bispinosa Foto: http://www.wetwebmedia.com/
Vissen zijn opmerkelijke wezens, en ze hebben zich uitzonderlijk goed aangepast aan het harde leven in de oceanen. Zelfs wanneer in groepen van vissen onvoldoende mannetjes of vrouwtjes beschikbaar zijn, kunnen veel soorten gewoon hun geslacht veranderen om voor de voortplanting te zorgen. De geslachtsverandering van alle vissoorten wordt gecontroleerd door sociale interacties. Een machtspositie, of ondergeschikte, is de trigger voor de geslachts hormoonspiegels, en zal leiden tot een verandering van geslacht in sommige situaties.
Bibliografie:
Wittenrich ML, Munday PL, Bi-directional sex change in coral reef fishes from the family Pseudochromidae: an experimental evaluation, Zoolog Sci. 2005 Jul;22(7):797-803
Sakai Y, Karino K, Kuwamura T, Nakashima Y, Maruo Y, Sexually dichromatic protogynous angelfish Centropyge ferrugata (Pomacanthidae) males can change back to females, Zoolog Sci. 2003, May;20(5):627-33.
Scholenvissen - Gobies
Scholenvissen - Gobies
Door Matt Wandell (vertaling en bewerking: Patrick Scholberg)
In een vorig artikel werd de Apogon parvulus behandeld en hier gaan we verder met twee andere mooie kleinoden die uitstekend geschikt zijn om zelfs nano-aquaria mee te bevolken. Hier gaan we wat dieper in op het gedrag en de huisvesting van de Trimma tevegae en de Aioliops megastigma.
Deze soorten komen uit totaal verschillende habitats en stellen dus heel andere eisen, de gemeenschappelijke kenmerken zijn de grootte of preciezer gezegd het gebrek eraan en het feit dat ze uitstekend geschikt zijn om met meerdere exemplaren tesamen gehouden te worden. Dus als je zo’n soort visje zoekt dan is dit artikel echt iets voor jou.
Trimma tevegae.

Foto: Tim Wong
Zoals nagenoeg alle Trimma’s blijft ook deze soort het zowat houden bij 2,5 cm voor een volwassen exemplaar en is dus uitermate geschikt voor zelfs kleine aquaria. Hij is gemakkelijk te houden en best wel sterk.
Wat deze soort van zijn soortgenoten onderscheidt is zijn gedrag en verdraagzaamheid. De meeste Trimma’s verschuilen zich tussen koralen of hangen onder rotsformaties en dat zowel op het rif als in het aquarium. Hierdoor zijn ze zeer moeilijk te bespeuren en moet je er echt naar op zoek gaan. Deze soort niet dus, hij staat als het ware mooi in de stroming in een meer verticale positie en probeert zo kleine partikels zooplankton naar binnen te werken.

Foto: Tim Wong
In tegenstelling tot veel andere Trimmasoorten die allen zeer hardnekkig hun eigen territorium verdedigen kan deze gobie makkelijk in een groepje van 6 à 8 exemplaren in een bakje van om en bij de 60 liter geplaatst worden en dan is dat een vrij realistische imitatie van de situatie op het rif.
Hoe ziet zijn natuurlijke habitat er uit? Wel, bouw in het aquarium een verticale of steile rifwand op en voorzie die van talrijke holen en inhammen waar ze zich in geval van nood in veiligheid kunnen brengen. De Trimma is meestal niet in de bovenste waterlagen te vinden waardoor we niet zo’n sterke verlichting moeten voorzien en we kunnen dus kiezen voor de wat hogere Kelvinwaarden waar de kleuren ook nog feller uit komen. Deze visjes kunnen zich echter zonder problemen aanpassen aan een hogere lichtintensiteit zolang we de overgang maar gradueel doorvoeren en hun de tijd gunnen.
Het voederregime voor deze visjes instellen is simpel zolang we maar vaak genoeg voeren, ze eten probleemloos diepvries-, droogvoer en pellets op zeer korte termijn alhoewel het soms nodig is hun in het begin nog wat levende artemia te voeren. Bij deze Gobie moeten we er ook rekening mee houden dat er ook flink gevoerd mag worden want door in het water op positie te blijven verbruikt hij beduidend meer energie dan de soorten die zich wegsteken tussen de koralen. Waar vaak andere Trimma’s in een bak gehuisvest kunnen worden zonder veel bijvoeren en van het kleine leven verder kunnen zul je snel merken dat voor deze kereltjes deze vlieger niet zo opgaat. Voorzie dus gerust 2 voerbeurten per dag of in geval van één voerbeurt smeer ze voldoende lang in de tijd uit zodat deze visjes zeker echt aan bod komen en niet verzwakken.
Geef dus genoeg voer, zorg voor een goede waterkwaliteit en zie af van vissen die deze kleinoden als lekkere hapjes beschouwen.
Als je aan deze punten de nodige aandacht besteedt dan leeft onze Trimma zowat 2 jaar in je aquarium.

Foto: Tim Wong
Er is eigenlijk maar één nadeel aan ons visje en dat is dat ze helaas lang niet overal te verkrijgen zijn, maar degene die moeite doet zal ze echter wel weten op de kop te tikken. Voor 2004 was dat een heel andere situatie maar de boom van de nano’s zorgt toch voor een veel betere beschikbaarheid van mooie Trimma’s en Eviota’s.
Aioliops megastigma

Deze gobie zie je zeker niet alledaags in de aquariumhandel maar te vinden zijn ze zeker wel, maar ja, je moet er moeite voor doen. Ze zijn nauw verwant aan de torpedogrondels maar met hun zowat 2 cm lengte zijn ze heel wat kleiner. Aangezien ze ook zeer slank zijn zorgt dit voor hun zeer specifieke eisen en kenmerken. Deze visjes bewegen zich nagenoeg continu door de waterkolom en gaan in tegen de stroming om zo de kleine zooplanktondeeltjes die hun tegemoet stromen naar binnen te werken.
Hun habitat is een bak met veel licht, een stevige stroming en een mix van zowel steen- als zachte koralen. In de natuur kennen ze een ruime verspreiding in de Stille Oceaan waar ze zich in groep ophouden boven koraalformaties.
In het aquarium moeten we er zeker in het begin aan denken onze stromingspompen te bedekken met een kous en een raster plaatsen voor de afloop blijkt geen overbodige luxe te zijn. Voorzie ook een flinke boord rondom je aquarium want deze kereltjes kunnen behoorlijke sprongen maken en het is niet de bedoeling ze ‘s morgens verdroogd op de grond terug te vinden.
( Noot: Deze opmerking geldt ook voor de Apogon parvulus die ik in een vorig artikel besproken heb; PERSOONLIJKE ERVARING van de vertaler die zijn eigen school van 27 exemplaren binnen het jaar zag slinken tot 10 stuks omwille van die reden ).
Bijna overbodig om te vermelden dat de medebewoners zorgvuldig uitgekozen moeten worden om deze visjes veilig in ons bakje te huisvesten. Grote brokkelsterren, Lysmata garnalen, zelfs heel wat koralen zijn in staat deze visjes te vangen en te verorberen en dan hebben we het niet eens over baarsjes, koraalwachters, Pseudochromis- of grotere Apogonsoorten die we er angstvallig van weg moeten houden. De gewone “opruimploeg” in een aquarium stelt echter geen probleem; heremietkreeftjes, zee-egels, slakken, mandarijnvisjes, pitvisjes… maar ook zeenaalden en andere kleine Gobies kunnen gerust toegevoegd worden.

Het heikele punt in de verzorging van deze visjes is hun snelle metabolisme en gezien de grootte van de voedseldeeltjes die ze kunnen bemachtigen zal dat in den beginne zeker geen sinecure zijn omdat ze aanvankelijk enkel op levend voer ingesteld zijn. Pas uitgekomen artemia kan dan soelaas brengen, iets ouder moet zeker verrijkt worden. Degene die beschikt over een cultuur van rotiferen, copepoden kan voor extra-afwisseling in hun voeding zorgen. Begin zeker ook tijdig te mengen met kwaliteitsstofvoeren die actueel op de markt voorhanden zijn.
Wat is nog een bijkomend nadeel? Hun levensduur schijnt om en bij het jaar te liggen en als we dan volwassen exemplaren bij de handelaar aantreffen dan is het moeilijk in te schatten hoe lang deze diertjes nog hebben om in ons aquarium verder door te brengen.

Referenties
Randall, J.E. and J.C. Delbeek. 2009. Comments on the extremes in longevity in fishes, with special reference to the Gobiidae. 447-454, 6 figs., 1 table. Proceedings of the California Academy of Science, Series 4, V60, Nos. 1-9.
De Roodvlek Kardinaalbaars Apogon parvulus
De verzorging van de Roodvlek Kardinaalbaars Apogon parvulus
Door Matt Wandell (vertaling, bewerking en foto's: Patrick Scholberg)
Vissen in een zeewateraquarium in een school houden brengt leven in een aquarium. Wat voor onze zoetwater collega’s heel vanzelfsprekend is, is een grote uitzondering in de zeewater aquaristiek. Sociaal gedrag kan tot uiting komen en zelfs voortplantingsrituelen kunnen tentoon gespreid worden. De mannetjes kleuren dan vaak fantastisch, in Amerika worden deze Supermales genoemd, je ziet dit als er voldoende vrouwtjes als stimulans dienen.
In een aquarium gaan zeewatervissen in eerste instantie een school vormen maar al heel vlug valt die uiteen omdat ze merken dat er geen echte bedreigingen zijn.

Nu is het gevaar dat uitgaat van rovers in de natuur slechts één van de redenen voor schoolvorming maar ook de kans op voedsel vergaren stijgt. Copepoden springen als ze nagezeten worden door haringen, als ze opgejaagd worden door een school zal zo’n sprong om aan de kaken van de ene haring te ontsnappen de copepod in het bereik van een andere haring positioneren. En studies wijzen uit dat haringen in de natuur deze afstand ook respecteren.
Voorts moeten we ook nog rekening houden in het zeewateraquarium dat heel wat vissen gedurende hun jeugd scholen vormen maar naarmate de groei vordert steeds exemplaren verdwijnen in het aquarium totdat slechts maximaal 2 exemplaren overschieten. We zien dit bij het vuurvisje, bij Chromis viridis.
Ook worden een aantal vissen veel te groot om een school in het aquarium te vormen, denk maar aan de doktersvissen en al zou je een zeer groot aquarium hebben dan nog is er ongekend veel agressiviteit te verwachten.
In grote aquaria kunnen Anthiassen zeer mooi in school gehouden worden mits we voldoende aandacht hebben voor goede waterwaardes doch dit mag nooit het argument zijn om de vissen minder te voeren, dan moet de school minder groot zijn.
Apogon parvulus (Roodvlek Kardinaalbaars)


Apogon parvulus zou in de natuur net geen 4cm groot worden, in onze aquaria halen ze ongeveer 2,5 cm. Het zilverkleurige lijf vertoont een blauwachtige glans en aan de staartwortel is een mooie rode vlek te zien. Door zijn gering formaat en vriendelijk gedrag is hij uitstekend geschikt voor het wat kleinere aquarium. Hierdoor stond hij bij heel wat liefhebbers op het verlanglijstje om zo een schooltje in het aquarium te huisvesten.
De eerste berichten waren echter alles behalve bemoedigend te noemen, uitval van 50% of meer was zeker geen uitzondering en enkele weken tot maanden later was geen enkel exemplaar meer in leven. Het was zo erg dat er vermoedens waren van een extreem korte levensduur wellicht minder dan één jaar. En al vlug gingen stemmen op om de visjes te laten waar ze vandaan kwamen, in de oceaan.
Niettegenstaande de negatieve berichten waagde de auteur in oktober 2009 zijn kans met een groepje van 18 visjes. Eéntje overleefde de aankomst niet en een ander stierf gedurende de quarantaine van 30 dagen. Twaalf van de zestien leven 14 maanden later nog wanneer deze tekst geschreven werd en zien er dan gezond en wel uit.
Wat is dan de reden dat deze vissen het zo ongewoon goed doen in vergelijking met de ervaringen van anderen?
Eind 2009 vernam de auteur van Kevin Kohen ( directeur van LiveAquaria ) : Apogon parvulus kent enorme sterftecijfers voornamelijk door de zeer gebrekkige behandeling na de vangst. Deze visjes moeten sterk gevoerd worden na de import om aan te sterken en rust krijgen eer ze verder verhandeld mogen worden.
We hebben grote zendingen binnengekregen zonder noemenswaardige uitval. Dit is enkel en alleen te verklaren door een gepaste behandeling nadien. We plaatsen ze enkel bij kleine rustige visjes en voeren ze zeer grondig, ze zijn dan ijzersterk.
Sinds we dat toepassen hebben we nagenoeg geen verlies meer.
Zoals bij zo vele andere aquariumvissen wordt de staat waar we de vissen in bekomen bepaald door de behandeling die de dieren bekomen hebben doorheen het hele transport tot hier. Dit kunnen we niet controleren. Waar we wel een invloed op hebben is hoe we zo’n dier bij aankomst behandelen. Maar ook WAAR we onze vissen kopen. COMMUNICEER ook of je tevreden bent of niet. Als meerdere hobbyisten hun ervaringen bij de winkelier meedelen is hij veel alerter en kan hij zijn leveranciers ook aanspreken. Het is zeer belangrijk dat we kwaliteitsvis kunnen aankopen die nog lang en gezond in ons aquarium verblijft.
Apogon parvulus in het Rifaquarium

Vooropgesteld dat we met gezonde visjes kunnen vertrekken is de parvulus helemaal niet zo moeilijk te houden in het rifaquarium. In de tropen vinden we ze in de bovenste waterlagen en is het zoutgehalte en de temperatuur van een doorsnee aquarium zeker geschikt. Qua lichtregime en stromingspatroon is wat we in ons aquarium doorgaans aanbieden in orde.
De enige echte uitdaging is ze het door ons aangeboden voedsel te laten eten en dat zeker een paar keer per dag. Gedurende de eerste twee weken moet dit visje echt stevig gevoerd worden met kleine ( levende ) artemia, watervlo, raderdiertjes, Cyclop-eez. Dit mag ondersteund worden met mysis van goede kwaliteit, garnaaltjes, viskuit, diepvrieskreefte-eitjes…. Dan kan langzaamaan droogvoer ingebouwd worden zodat je ze meermaals per dag met de voederautomaat kan blijven voeren. Een goed rijp aquarium en/of refugium kan een belangrijke bijdrage leveren.
Gezien hun gering formaat is een aangepast visbestand nodig anders is de parvulus zelf een voedseldiertje. Let dus zeker op met baarsjes, koraalklimmers en alle vissen die groter en ruwer in de omgang zijn. Zelf is de parvulus geen bedreiging voor andere levende have behalve wanneer we uitgegroeide parvulussen een zeer kleine goby als gezelschap geven of er een kleine Thor amboinensis bij plaatsen.
Als we deze visjes gepast huisvesten, goed voeren kan dit geregeld tot broedsels leiden. De mannetjes zijn muilbroeders en houden de eitjes in de muil tot ze uitkomen. Hierdoor kunnen we ook de mannetjes onderscheiden. De larven zijn veel kleiner dan van andere Apogons en daardoor ook heel wat moeilijker groot te brengen waardoor hierover ook geen berichten bekend zijn.
In geval van bijkomende vragen kan de auteur bereikt worden op Dit e-mailadres wordt beveiligd tegen spambots. JavaScript dient ingeschakeld te zijn om het te bekijken. wel in het Engels uiteraard.
Evolutie van de kopmorfologie van Syngnathidae
Evolutie van de gespecialiseerde kopmorfologie van Syngnatidae
Dit is een samenvatting van het doctoraatsproefschrift van Heleen Leysen. 2011. Evolutionary morphology of the extremely specialized feeding apparatus in seahorses and pipefishes (Syngnathidae). Unpublished doctoral thesis. Ghent University. zie http://www.fun-morph.ugent.be/ en meerbepaald http://www.fun-morph.ugent.be/Projects/Seahorses/Seahorses.htm. Het volledig Engelstalig proefschrift kan je lezen op http://www.evomorph.ugent.be/Publications/Publ80.pdf
Dit doctoraatsproefschrift behandelt de evolutie van de gespecialiseerde kopmorfologie in Syngnathidae, een familie binnen de beenvissen waartoe zeenaalden, zeepaardjes, zeedraken en ‘pipehorses’ behoren. Een gedetailleerde morfologische beschrijving van het voedselopnameapparaat bij enkele syngnathe vertegenwoordigers vormt de kern van dit onderzoek. Daarnaast worden andere aspecten zoals morfologische variatie binnen de familie, werking van het trofisch systeem en mechanische belasting tijdens de voedselopname uitvoerig bestudeerd. De volgende samenvatting geeft een duidelijk overzicht van de belangrijkste resultaten en conclusies per hoofdstuk en biedt een antwoord op de onderzoeksvragen zoals geformuleerd in hoofdstuk 1.3.2.

In het eerste hoofdstuk worden enkele algemene begrippen zoals ‘evolutie’, ‘natuurlijke selectie’ en ‘specialisatie’ toegelicht. Daarop volgt een inleiding tot de biologie van Syngnathidae, waarbij de taxonomische positie, voortplanting, complexe broedzorg en uitwendige morfologie aan bod komen. Vervolgens wordt de wetenschappelijke probleemstelling geschetst. Het voedselopname-appararaat is in deze familie extreem gespecialiseerd. Alle vertegenwoordigers hebben een buisvormige snuit met kleine, eindstandige kaken. De lengte en diameter van de snuit is echter variabel over verschillende soorten. Voedselopname (‘pivot feeding’) gebeurt door middel van koprotatie waardoor de snuittip naar de prooi gebracht wordt, gevolgd door een volumetoename in de buccale holte.
Deze expansie creëert een onderdruk waardoor een waterstroom in de richting van de mond ontstaat. Het vangen van een prooi gebeurt heel snel (minder dan 6 ms), maar voedselopname met behulp van een lange, smalle snuit houdt ook een aantal hydrodynamische beperkingen in (zoals een hoge weerstand bij het zuigen van water door een dunne buis en een groot inertiemoment tijdens rotatie van een kop met lange snuit). Deze paradox maakt het voedselopname-apparaat bij Sygnathidae tot een zeer interessant onderzoeksobject. De doelstelling van dit doctoraatsonderzoek is dan ook het ontrafelen van het evolutionaire patroon dat aan de basis ligt van deze extreme morfologische en functionele specialisatie.
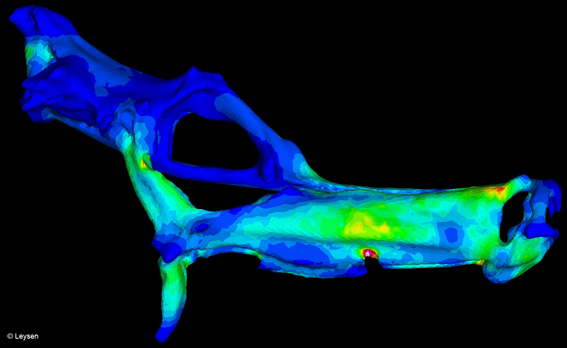
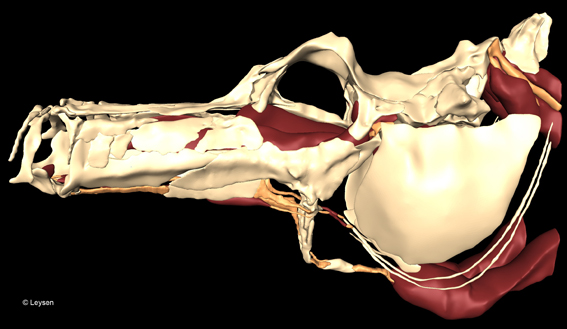
Hoofdstuk twee geeft een overzicht van het bestudeerde materiaal en de toegepaste methodes. Eerst wordt een opsomming gegeven van de gekozen soorten met vermelding van de herkomst van de specimens en waarvoor ze gebruikt werden. In een tweede deel worden de technieken die doorheen het onderzoek toegepast werden, uitgebreid uitgelegd. Gedetailleerde protocols voor het ophelderen en kleuren van specimens en voor het maken van histologische coupes worden gegeven, alsook de gehanteerde methode bij het meten, uitvoeren van dissecties, nemen van CT-scans en maken van grafische 3D-reconstructies. Iets meer aandacht wordt besteed aan de geometrisch morfometrische en de eindige elementen (finite element) analyses. Tenslotte volgt in de laatste paragraaf een verduidelijking van een aantal controversiële begrippen.
Het derde hoofdstuk handelt over de morfologie van het voedselopnameapparaat bij Syngnathidae en is onderverdeeld in twee delen. In het eerste deel wordt de bouw van het cranium van een zeenaald (Syngnathus rostellatus) en van een zeepaard (Hippocampus capensis) in een evolutieve context geplaatst door vergelijking met de kopmorfologie van een stekelbaars (Gasterosteus aculeatus). Ook de morfologie van de juveniele kop rond het tijdstip van vrijkomen uit de buidel, komt aan bod. De snuitverlenging van de kraakbenige schedel situeert zich ter hoogte van de ethmoid regio en de ethmoidplaat, het hyosymplecticum en het basihyale zijn verlengd. In de juvenielen zijn alle craniale beenderen, met uitzondering van de circumorbitalia en enkele operculaire beenderen, al gevormd, zij het enkel als een zeer dun laagje.
Deze grote gelijkenis met de adulte kop is waarschijnlijk te wijten aan de lange broedperiode en uitgebreide ouderlijke zorg. Hierdoor kan het vrijlaten van de jongen uit de buidel uitgesteld worden tot ze een vergevorderd ontwikkelingsstadium bereikt hebben. Verder tonen de resultaten aan dat de adulte snuit gevormd wordt door een verlenging van het vomer, mesethmoid, quadratum, metapterygoid, symplecticum en preoperculare. Enkele morfologische aspecten die als evolutionaire adaptaties aan de gespecialiseerde voedselopname kunnen beschouwd worden, zijn de gereduceerde en tandloze maxillaire beenderen, de stevige verbinding tussen de verschillende beenderen van de onderkaak, het robuuste kieuwdeksel en het zadelvormig gewricht tussen interhyale en ceratohyale posterior.
In het tweede deel van dit hoofdstuk ligt de nadruk op het effect van snuitverlenging binnen de familie, zowel op osteologisch als myologisch vlak. Een gedetailleerde morfologische studie van de kop van zowel een lang-snuitige zeepaard (H. reidi) als een zeenaald met extreem lange snuit (Doryrhamphys dactyliophorus), in vergelijking met een aantal syngnathen met korte en intermediaire snuit (H. zosterae, H. abdominalis, Corythoichthys intestinalis en D. melanopleura), bevestigt de resultaten uit het voorgaande deel. Bovendien worden enkele gespecialiseerde spier-, pees-en ligamentconfiguraties aan het licht gebracht. Zo is de sternohyoideus spier heel goed ontwikkeld bij zeepaardjes, wordt de protractor hyoidei spier bij enkele syngnathen omgeven door het mandibulo-hyoid ligament en wordt in de epaxiale pezen van een aantal zeenaalden een lang, staafvormig sesamoid been gevonden. Deze waarnemingen suggereren de aanwezigheid van verschillende vermogen versterkende strategieën voor koprotatie tijdens de voedselopname. Het vrijlaten van elastische energie, opgeslagen in de epaxiale en hypaxiale pezen, lijkt begunstigd in zeenaalden terwijl spierkracht verkozen wordt in zeepaardjes. Hoewel het syngnathe musculoskeletale systeem uit een aantal elementen bestaat die gedeeld worden door alle vertegenwoordigers, zijn er dus toch een aantal opmerkelijke morfologische verschillen tussen soorten. Slechts weinig van deze verschillen kunnen echter gerelateerd worden aan variatie in relatieve snuitlengte.
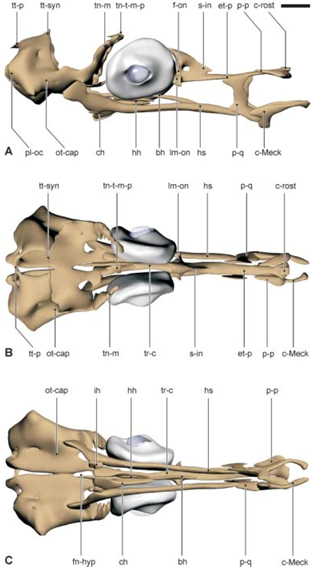
Fig 4. 3D reconstructie van het juveniele chondrocranium van Syngnathus rostellatus UGMD175388 (13.1 mm SL)
Aansluitend bij het voorgaande, wordt in hoofdstuk vier de morfologische variatie binnen de familie gekwantificeerd door middel van een geometrisch morfometrische analyse bij een groot aantal syngnathe vertegenwoordigers. Via deze methode kan vastgesteld worden dat de belangrijkste verschillen in kopmorfologie tussen zeenaalden en zeepaardjes gerelaterd zijn aan veranderingen in snuit-en kopdimensie, de positie van de pectorale vin en de hoogte van het kieuwdeksel. Deze variatie heeft waarschijnlijk te maken met de oriëntatie van de lichaamsas ten opzichte van de kop (lichaam in het verlengde of onder een hoek met de kop) en aan de kinematica van de voedselopname (lange snuit en beperkte koprotatie of korte snuit en veel rotatie).
Gezien de hoge graad van specialisatie in het voedselopname-apparaat bij Syngnathidae zou een kleine afwijking in de interactie tussen de verschillende elementen in de kop al tot een verminderde prestatie kunnen leiden. Dit doet vermoeden dat elke soort binnen de familie gekenmerkt wordt door een beperkte intraspecifieke morfologische variatie. Vooral de morfologische plasticiteit van soorten met een lange snuit, die als meer gespecialiseerd beschouwd worden dan kort-snuitige soorten, zal gereduceerd zijn. De preliminaire resultaten van deze studie bevestigen onze verwachtingen.
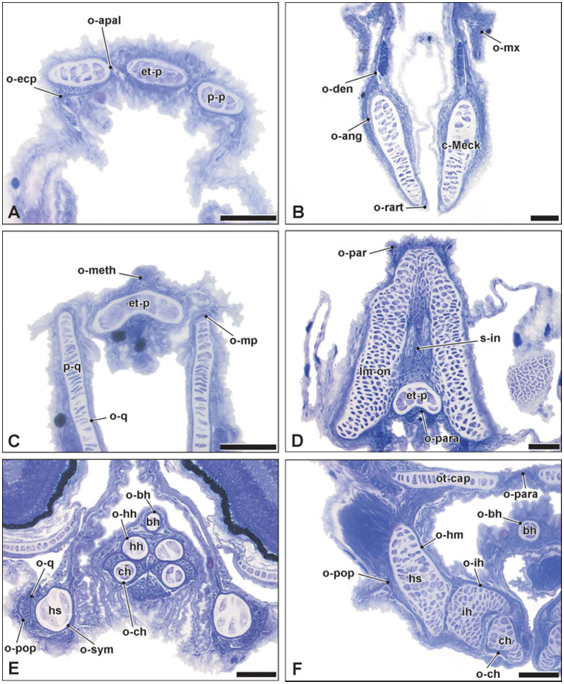
Fig 5 Histologische doorsnede van het juveniele cranium van Syngnathus rostellatus UGMD175388 (13.1 mm SL)
Tenslotte wordt onderzocht of er nog belangrijke ontogenetische transformaties optreden bij juveniele zeepaardjes (H. reidi) na het vrijkomen uit de buidel. Graduele vormverschuivingen, zoals het dorsoventraal vernauwen van de snuit en kop en de heroriëntatie van het preoperculair been, kenmerken de juveniele ontwikkeling. Maar, de periode van ingrijpende veranderingen lijkt reeds doorlopen te zijn in de buidel
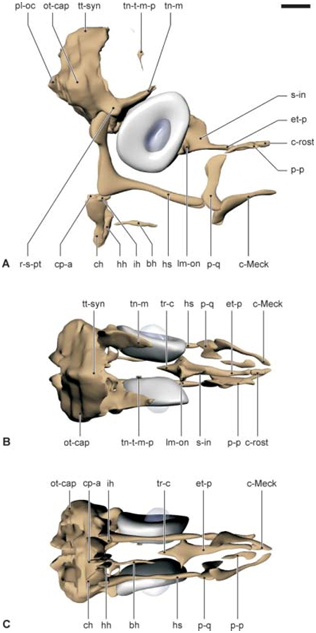
Fig. 6. 3D reconstructie van het juveniele chondrocranium van Hippocampus capensis UGMD175394 (12.8 mm SL)
In het vijfde hoofdstuk komen enkele functionele aspecten van het gespecialiseerde voedselopname-apparaat aan bod. Het eerste deelonderzoek werd uitgevoerd in samenwerking met Gert Roos en Sam van Wassenbergh (Universiteit Antwerpen) en evalueert een eerder beschreven planair vierstangensysteem (Muller, 1987) dat rotatie van het hyoid koppelt aan neurocraniale elevatie. Zowel de morfologische beschrijving van alle stangen en gewrichten betrokken bij het systeem, als de kinematische opnames van de voedselopname bij een zeepaardje (H. reidi) tonen de ongelooflijke specialisatie van het trofisch apparaat aan.

Hippocampus reidi Foto: Hystrix, uit het aquarium van Piet Hectors
Niet alleen verschilt de syngnathe ‘pivot feeding’ van zuigvoeding bij een gegeneralizeerde teleost in snelheid (duur van een prooiopname is minder dan 6 ms), maar ook de volgorde van beweging is anders. De expansiefase begint met hyoidrotatie en niet met mondopening zoals gebruikelijk (rostrocaudale golf). Bovendien leveren de resultaten een duidelijk bewijs voor de koppeling tussen hyoid en neurocranium. Er wordt echter een discrepantie tussen de voorspelde bewegingen (op basis van het vierstangenmodel) en de waargenomen bewegingen (op basis van de videoopnames) aangetoond.
Dit is hoogst waarschijnlijk te wijten aan het modelleren van de stang bestaande uit het urohyale en de sternohyoideus spier als onveranderlijk in lengte, terwijl deze verkort bij spiercontractie. In het tweede deel wordt onderzocht waar in de schedel de meeste mechanische stress optreedt als gevolg van de grote drukveranderingen tijdens de voedselopname. Hiervoor wordt een eindige elementen analyse uitgevoerd op de gemodelleerde schedels van drie zeepaard-en drie zeenaaldsoorten met variërende relatieve snuitlengte (H. reidi, H. abdominalis, H. zosterae, D. dactyliophorus, S. rostellatus en D. melanopleura). In overeenstemming met de verwachtingen (uitgezonderd in D. dactyliophorus) worden stresspieken waargenomen ter hoogte van articulaties en kraakbenige zones in de gemodelleerde schedels. Deze concentraties aan stress zullen in de werkelijke schedels veel lager liggen aangezien zowel kraakbeen als articulerende elementen verondersteld worden stress goed te weerstaan.
Een tweede doel is nagaan of er een verschil is in stressdistributiepatroon tussen kort-en langsnuitige soorten. De hypothese is dat soorten met een lange snuit grotere drukverschillen moeten creeëren, hogere stress moeten kunnen weerstaan en dus bepaalde morfologische adaptaties zullen hebben die dit toelaten. Wanneer kort-en langsnuitige modellen blootgesteld worden aan eenzelfde druk, zullen de modellen met een lange snuit de minste stress ervaren. Deze hypothese wordt niet bevestigd door de resultaten, maar de geometrie van de beenderen in de snuit (dikte van het been, mate van overlap tussen de beenderen, enz.) heeft een duidelijke invloed op de stress distributie.
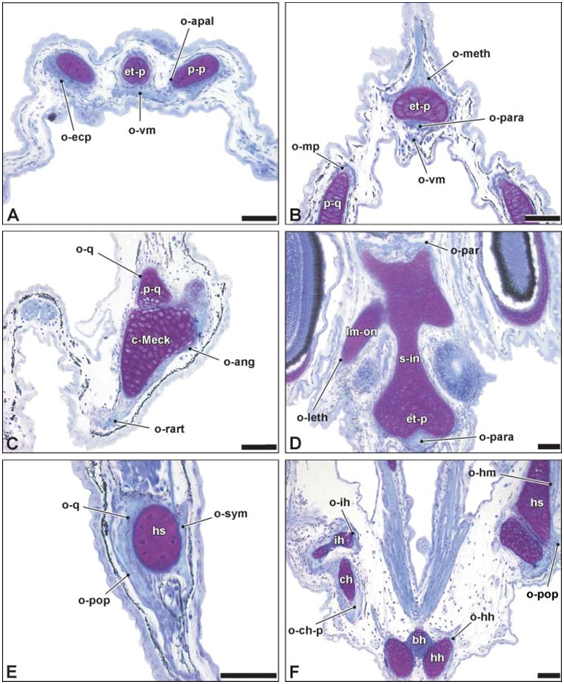
Fig. 7. Histologische doorsnede van het juvenile cranium van Hippocampus capensis UGMD175394 (12.8 mm SL)
In hoofdstuk zes worden de behaalde resultaten bediscussieerd in een ontogenetische, functionele en evolutionaire context, gevolgd door een algemene conclusie. Ten eerste komt de bijzondere ontwikkeling bij Syngnathidae aan bod. De broedzorg duurt lang en is complex; het mannetje hecht de eitjes en embryos aan het lichaam vast of bewaart ze in een afgesloten buidel, zelfs nadat ze uit het ei gekomen zijn. De mate van ouderlijke investering neemt toe met toenemende complexiteit van de broedstructuur. Zelfs de meest basale syngnathen bieden de jongen bescherming tegen predatie, meer geëvolueerde soorten zorgen ook voor osmoregulatie, voeding en zuurstoftoevoer. Door de verlenging van de broedperiode wordt de start van exogene voedselopname uitgesteld en is er meer tijd voor de ontwikkeling van de jongen. Pasgeboren syngnathen zijn dan ook bijna volledig ontwikkeld (de larvale fase en bijhorende metamorfose zijn afwezig of vinden plaats in de buidel) en juvenielen kunnen zelfstandig prooien vangen door middel van de speciale ‘pivot feeding’. Samenvattend kan gesteld worden dat de ingrijpende broedzorg waarschijnlijk de evolutie van de gespecialiseerde syngnathe kopmorfologie in de hand heeft gewerkt. In het tweede deel van de discussie wordt de nadruk gelegd op de werking van de syngnathe kop. De belangrijkste structurele en functionele modificaties in vergelijking met het trofische apparaat van een gegeneraliseerde teleost worden herhaald. Daarna volgt een gedetailleerde beschrijving van de beweging van en interactie tussen de afzonderlijke elementen in de kop tijdens de voedselopname.
Tenslotte worden de voor-en nadelen gerelateerd aan snuitverlenging opgesomd. Hieruit blijkt dat de morfologische variatie in termen van snuitdimensie in de familie waarschijnlijk het gevolg is van een trade-off. Voor de optimalisatie van koprotatie is een lange snuit voordelig, terwijl voor het genereren van een grote zuigkracht een korte snuit gunstiger is. De grote morfologische diversiteit binnen de Syngnathidae zou zo verklaard kunnen worden. Het derde deel behandelt de resultaten in een fylogenetisch en evolutionair kader. Er wordt eerst kort beargumenteerd waarom vergelijking van de syngnathe kop met die van de driedoornige stekelbaars (Gasterosteus aculeatus) nog steeds nuttig is, ook als ze niet elkaars zustertaxa blijken te zijn. Vervolgens worden de evolutionaire trends besproken die tot de zeepaardmorfologie (d.w.z. de grijpstaart, de gekantelde kop en de verticale lichaamshouding) zouden hebben kunnen leiden.
Het genus Idiotropiscis (‘pygmy pipehorses’) wordt beschouwd als de nog levende transitievorm tussen zeenaalden en zeepaardjes. De divergentie van pipehorses en zeepaarden zou bevorderd zijn door de opmars van zeegrashabitats die meer bescherming bieden aan organismen met een rechte lichaamsas. Ook de biomechanica van de voedselopname zou een rol gespeeld kunnen hebben in de evolutie van zeepaardjes. De kanteling van de kop ten opzichte van de lichaamsas zou koprotatie kunnen bevoordelen en de grijpstaart laat een strekking van het lichaam in de richting van de prooi toe. Deze voorwaartse beweging van de mond zou de tijd die nodig is om de prooi te bereiken reduceren, wat precies het effect van snuitverlenging is. Als een grote koprotatie in combinatie met een voorwaartse beweging in de richting van de prooi hetzelfde selectief voordeel biedt als een lange snuit, zou dat een verklaring kunnen zijn voor het ontbreken van zeepaardjes met uitzonderlijk lange snuit. Tenslotte worden deze bevindingen kort en bondig samengevat in de algemene conclusie.
Inloggen
Registreren
Foto van de maand

Blauwe mandarijnpitvis Synchiropus splendidus
Foto: Sander Hemmes